You might also like
- El Poder de Los HabitosDocument1 pageEl Poder de Los HabitosJose Jiménez50% (6)
- Estado de Cuenta Junio 2023Document6 pagesEstado de Cuenta Junio 2023danielNo ratings yet
- Germanica Nueva MedicinaDocument11 pagesGermanica Nueva Medicinafelipe4alfaro4salas100% (1)
- LA PERSECUCION DE LA OVEJA SALVAJE - Ruben - MDocument28 pagesLA PERSECUCION DE LA OVEJA SALVAJE - Ruben - MRubenMarinNo ratings yet
- Hematologia ForenseDocument46 pagesHematologia ForenseManuel Hernandez Luis100% (1)
- Pin de Diagnostico y de Computadira Toyota 4EDocument5 pagesPin de Diagnostico y de Computadira Toyota 4ETecn Jhuliño Andahua Lopez100% (1)
- Parametros Fisico-Quimicos en AcuiculturaDocument4 pagesParametros Fisico-Quimicos en AcuiculturaRoberto Carlos Vasquéz CastañedaNo ratings yet
- Informe Ecosistemas AcuaticosDocument9 pagesInforme Ecosistemas AcuaticosLuzFabiolaAlapeOrtizNo ratings yet
- 209929513.1 Introducción A La Ictiología PDFDocument13 pages209929513.1 Introducción A La Ictiología PDFEdgar Galeano100% (1)
- SESION 3 - Parámetros de Calidad Del AguaDocument35 pagesSESION 3 - Parámetros de Calidad Del AguahenryllecllishNo ratings yet
- Historia de La Terapia Familiar - CarlosgavanchoDocument10 pagesHistoria de La Terapia Familiar - CarlosgavanchoCarlos Gavancho0% (1)
- Clasificacion Termica de Los LagosDocument18 pagesClasificacion Termica de Los LagosMilagritos Gianelly Sanchez Zarate100% (7)
- Ecosistemas de Aguas Dulces y Aguas SaladasDocument17 pagesEcosistemas de Aguas Dulces y Aguas SaladasFernando José Zapata BardalesNo ratings yet
- Manual de IctiologiaDocument83 pagesManual de IctiologiaLumikoM.TasaycoNo ratings yet
- Filosofia de La Historia para La Educacion de La Humanidad (Von Herder Johann Gottfried)Document143 pagesFilosofia de La Historia para La Educacion de La Humanidad (Von Herder Johann Gottfried)reginaldo aliçandro Bordin100% (1)
- Clase 5 - Estratificacion TermicaDocument41 pagesClase 5 - Estratificacion TermicaDani Danitho FernandezNo ratings yet
- Especies Marinas Invertebrados. Didacticas de Las CienciasDocument9 pagesEspecies Marinas Invertebrados. Didacticas de Las Cienciasana camila castrillonNo ratings yet
- Zonacion Sistemas Loticos, LenticosDocument48 pagesZonacion Sistemas Loticos, LenticosGon Negro Alfred100% (1)
- MACROINVERTEBRADOSDocument26 pagesMACROINVERTEBRADOSWilder Doneth Oblitas AbantoNo ratings yet
- Rio ContinuoDocument10 pagesRio ContinuoSimone Cárdenas YelaNo ratings yet
- Plan de Manejo Cacajao CalvusDocument21 pagesPlan de Manejo Cacajao CalvusYomar malac Noriega delgadoNo ratings yet
- Delimitacion de EcosistemaDocument21 pagesDelimitacion de Ecosistemaluz112No ratings yet
- Metabolismo de Ecosistemas AcuaticosDocument66 pagesMetabolismo de Ecosistemas AcuaticosJohn Graham40% (5)
- Estratificación Térmica de Un LagoDocument2 pagesEstratificación Térmica de Un LagoAnonymous S0BZX8No ratings yet
- Lago - LimnologiaDocument9 pagesLago - LimnologiaMercedes Villa0% (1)
- Informe Ecologia 2 Parametros FisicoquimicosDocument12 pagesInforme Ecologia 2 Parametros FisicoquimicosANGIE VANESSA MENDOZA ARRIETA (:-)No ratings yet
- Clasificación de Los LagosDocument3 pagesClasificación de Los LagosAndres Morera Ortiz100% (1)
- Funcione de La Piel de Los PecesDocument4 pagesFuncione de La Piel de Los PecesCelestinaNo ratings yet
- Anatomía de Los PecesDocument13 pagesAnatomía de Los Pecesאנטוניו סנטאןNo ratings yet
- INFORME - EcosistemasDocument15 pagesINFORME - EcosistemasNicole MendozaNo ratings yet
- Locomoción y Migraciones II - ICTIOLOGIADocument17 pagesLocomoción y Migraciones II - ICTIOLOGIAFiorela RuizNo ratings yet
- Estudio y Analisis de Macroinvertebrados Acuaticos Como Bioindicadores en La Cuenca Del Rio Combeima-1Document32 pagesEstudio y Analisis de Macroinvertebrados Acuaticos Como Bioindicadores en La Cuenca Del Rio Combeima-1Marco Gonzalez100% (1)
- GUIA LimnologiaDocument12 pagesGUIA LimnologiajorelljackNo ratings yet
- Factores AbioticosDocument3 pagesFactores AbioticosMargarita Maria Gaitan VeraNo ratings yet
- Limnología y EutrofizaciónDocument16 pagesLimnología y EutrofizaciónCarlos Benjamin EspinosaNo ratings yet
- Sistema Circulatorio PecesDocument11 pagesSistema Circulatorio PecesJoseUreyNo ratings yet
- Capitulo 1 Introduccion Definiciones Diagrama de RawsonDocument26 pagesCapitulo 1 Introduccion Definiciones Diagrama de RawsonYazzmin VasquezNo ratings yet
- Ecosistemaacuatico 130211113018 Phpapp01Document13 pagesEcosistemaacuatico 130211113018 Phpapp01Carlos MéndezNo ratings yet
- Choro FinalDocument17 pagesChoro FinalRicardo Segura FloresNo ratings yet
- Biometria de Los Peces ImprimirDocument8 pagesBiometria de Los Peces ImprimirFreddyNo ratings yet
- Aplicasion de Rotiferos en LarviculturaDocument3 pagesAplicasion de Rotiferos en Larviculturalordj_10No ratings yet
- Técnicas de Muestreo y Análisis Cuantitativo y Cualitativo Del FitoplanctonDocument14 pagesTécnicas de Muestreo y Análisis Cuantitativo y Cualitativo Del FitoplanctonCesar Eduardo Manrique GraosNo ratings yet
- Ensayo EutrofizaciónDocument2 pagesEnsayo EutrofizaciónWilliam CedeñoNo ratings yet
- Perfil de Tesis-Plantas Acuaticas - Manuel - SGDocument26 pagesPerfil de Tesis-Plantas Acuaticas - Manuel - SGManuel Jesus Marca ZevallosNo ratings yet
- Qué Es El Monitoreo BiológicoDocument2 pagesQué Es El Monitoreo BiológicoWily WillsNo ratings yet
- Fijación y Preservación de Anfibios y ReptilesDocument1 pageFijación y Preservación de Anfibios y ReptilesHugoNo ratings yet
- Origen y Evolucion de Los Mamiferos PDFDocument5 pagesOrigen y Evolucion de Los Mamiferos PDFbetagam12777No ratings yet
- Aguas Lénticas o EstancadasDocument22 pagesAguas Lénticas o EstancadasFrancisco Javier Benitez Lira88% (8)
- Ecologia de Las ComunidadesDocument12 pagesEcologia de Las ComunidadesHuereqq LambayeqNo ratings yet
- Algas de Agua DulceDocument3 pagesAlgas de Agua DulceRx SangaNo ratings yet
- Informe - Caracterización de Lago YarinacochaDocument18 pagesInforme - Caracterización de Lago YarinacochaCarlos Torres Cueva0% (2)
- Plantas FlotantesDocument9 pagesPlantas Flotantesmarcelo1315No ratings yet
- Cuenca Hidrográfica de AmazonasDocument5 pagesCuenca Hidrográfica de AmazonasMarisolNo ratings yet
- Tema 6 La Biosfera PDFDocument78 pagesTema 6 La Biosfera PDFcasaparra106No ratings yet
- Filo Nematodos (Nemata) B&BDocument16 pagesFilo Nematodos (Nemata) B&BJosber FerrerNo ratings yet
- Comunidades BióticasDocument3 pagesComunidades BióticasLuzma ChigüichónNo ratings yet
- Elementos de EcologiaDocument41 pagesElementos de EcologiaMoises TordoyaNo ratings yet
- RotiferosDocument7 pagesRotiferosIsrael Villar0% (1)
- Marcacion de PecesDocument20 pagesMarcacion de PecesZabdiel Ramirez HuansiNo ratings yet
- Flora y FaunaDocument95 pagesFlora y Faunaricardo floresNo ratings yet
- Caracterización Cuenca Río AyampeDocument86 pagesCaracterización Cuenca Río AyampeLuz Del Alba Meza Cruz100% (1)
- Pesca Con Linea de ManoDocument2 pagesPesca Con Linea de ManoCarola Ortiz MogollonNo ratings yet
- Adaptaciones A La Selva TropicalDocument96 pagesAdaptaciones A La Selva TropicalFredy LeonardoNo ratings yet
- ChrysophyceaeDocument11 pagesChrysophyceaesilemNo ratings yet
- Macrofitas de Agua DulceDocument18 pagesMacrofitas de Agua DulceCristhianBorisGutierrezQuispeNo ratings yet
- Ecosistemas. FuncionamientoDocument4 pagesEcosistemas. FuncionamientoStefaniSalazarGaonaNo ratings yet
- Ecología FluvialDocument13 pagesEcología FluvialJavier RiquelmeNo ratings yet
- Mauricio Daniel Coello NavarroDocument10 pagesMauricio Daniel Coello NavarroMine CruzNo ratings yet
- 26 - 85 - Punta LeonDocument2 pages26 - 85 - Punta LeonBabuflyNo ratings yet
- 26 - 72 - Punta TomboDocument2 pages26 - 72 - Punta TomboBabuflyNo ratings yet
- 26 - 67 - Punta LomaDocument2 pages26 - 67 - Punta LomaBabuflyNo ratings yet
- 26 - 73 - Cabo Dos BahiasDocument4 pages26 - 73 - Cabo Dos BahiasBabuflyNo ratings yet
- 26 - 06 - Piedra ParadaDocument2 pages26 - 06 - Piedra ParadaBabuflyNo ratings yet
- Ley 2580-85 - Modifica Ley 2161 (Punta Del Marquéz)Document1 pageLey 2580-85 - Modifica Ley 2161 (Punta Del Marquéz)BabuflyNo ratings yet
- Manejo de Recursos CulturalesDocument10 pagesManejo de Recursos CulturalesBabuflyNo ratings yet
- Entrega Reseña - Consigna para La Escritura de La Reseña. Coe 1. Gr.33 2021-2Document7 pagesEntrega Reseña - Consigna para La Escritura de La Reseña. Coe 1. Gr.33 2021-2R VNo ratings yet
- LeyesDocument14 pagesLeyesCameronNo ratings yet
- Cuestionario#2 Paula AlmanzaDocument9 pagesCuestionario#2 Paula AlmanzaPaula AlmanzaNo ratings yet
- GC-FT-188 Ficha Tecnica de Producto Terminado Rastrillo JardinDocument1 pageGC-FT-188 Ficha Tecnica de Producto Terminado Rastrillo JardinMarlon Esteban MARQUEZ LOPEZNo ratings yet
- Introduccion Fisioterapia CardiopulmonarDocument3 pagesIntroduccion Fisioterapia CardiopulmonarDaniela HernándezNo ratings yet
- Facultad Ciencias Empresariales Escuela Académica de ContabilidadDocument14 pagesFacultad Ciencias Empresariales Escuela Académica de ContabilidadIvanNo ratings yet
- Los Juegos de Crianza y El Holding Corporal Con RDocument17 pagesLos Juegos de Crianza y El Holding Corporal Con Rlupe ferreyraNo ratings yet
- Laboratorio 7Document5 pagesLaboratorio 7Laura CastroNo ratings yet
- Tipos de Anomalías GeoquímicasDocument8 pagesTipos de Anomalías GeoquímicasGabriela solis vazquezNo ratings yet
- ENVIO 2021 - InicialDocument5 pagesENVIO 2021 - InicialKIESER DE LA PEÑA GASPARNo ratings yet
- Cuadro Sinoptico de Planeacion DidacticaDocument1 pageCuadro Sinoptico de Planeacion DidacticaFrancisco Jahir HernandezNo ratings yet
- Obra de Teatro No Se Culpe A NadieDocument22 pagesObra de Teatro No Se Culpe A NadieCésar Huerta100% (2)
- Actron Autoscaner CP9575 PDFDocument16 pagesActron Autoscaner CP9575 PDFMelvin R. CisnerosNo ratings yet
- Siembra de Muestras Microbiológicas CASR - AspxDocument13 pagesSiembra de Muestras Microbiológicas CASR - AspxJaviera Viviana Véliz SilvaNo ratings yet
- Periodismo de Investigación 3 ActividadDocument8 pagesPeriodismo de Investigación 3 ActividadAna Ruiz ChirinoNo ratings yet
- Instrucciones Tm.Document27 pagesInstrucciones Tm.BereniceNo ratings yet
- Test OTISDocument10 pagesTest OTISMarco BravoNo ratings yet
- Claus Sluter - WikipediaDocument3 pagesClaus Sluter - WikipediaAnthony CerdasNo ratings yet
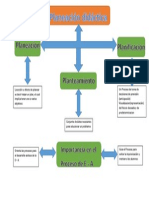
- Planeamiento Estrategico (5 Ciclo)Document10 pagesPlaneamiento Estrategico (5 Ciclo)CoQa Gu GullianNo ratings yet
- Organelos Citoplasmaticos 2022Document4 pagesOrganelos Citoplasmaticos 2022OlivSanMaria100% (1)
- ErisipelaDocument2 pagesErisipelaVandalicosterNo ratings yet
- Taz PFC 2015 129Document105 pagesTaz PFC 2015 129Navarro Viera Shary JazmínNo ratings yet