Professional Documents
Culture Documents
Módulo Reproducción Animal
Uploaded by
Alecsander RomeroCopyright
Available Formats
Share this document
Did you find this document useful?
Is this content inappropriate?
Report this DocumentCopyright:
Available Formats
Módulo Reproducción Animal
Uploaded by
Alecsander RomeroCopyright:
Available Formats
Universidad Estatal de Bolvar (FAC. CC.
AGROP) MVZ
UNIVERSIDAD ESTATAL DE BOLIVAR
FACULTAD DE CIENCIAS AGROPECUARIAS
RECURSOS NATURALES Y DEL AMBIENTE
ESCUELA DE MEDICINA VETERINARIA
Y ZOOTECNIA
MODULO
REPRODUCCIN ANIMAL
Catedrtica:
Dr. MVZ. ngela Caldern Tobar
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
NDICE
1. BASES MORFOLGICAS DE LA REPRODUCCIN/15
La pelvis como conducto/15
Eje pelviano/17
Diafragma plvico/19
Dimetros del estrecho anterior de la pelvis/19
Pelvis de las distintas especies domsticas/ 20
Dolicoplvica/20
Mesatiplvica/22
Platiplvica/24
Pelvimetra/ 24
Distintos tipos de tero/ 27
tero simple/27
tero bicornual/28
tero dividido o bipartido/ 30
tero doble/30
Variaciones de la anatoma de los rganos genitales/ 32
rganos genitales de la vaca/ 32
rganos genitales de la yegua/ 38
rganos genitales de los pequeos rumiantes/42
rganos genitales de la cerda/ 43
rganos genitales de la perra/ 45
Situacin intravital de los rganos genitales/ 48
Situacin intravital de los rganos genitales de la vaca/48
Situacin intravital de los rganos genitales de la yegua/ 50
Referencias bibliogrficas/ 52
2. FUNCIONES REPRODUCTIVAS/53
Ciclo reproductivo/ 53
Etapa pre-reproductiva/54
Etapa reproductiva/54
Etapa pos-reproductiva/54
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
Pubertad/ 55
Aspectos metablicos en las novillas asociados al sistema de alimentacin/57
Perfiles hormonales/59
Variaciones en otras especies/ 60
Ciclo estral/ 61
Duracin del ciclo estral/ 62
Fases del ciclo estral/62
Manifestaciones clnicas del celo en la vaca/ 64
Sintomatologa clnica del metestro. 66
Sintomatologa clnica del diestro/ 67
Modificaciones estructurales del tracto genital/ 71
Ciclo ovrico/71
Ciclo tubrico/84
Ciclo uterino/85
Ciclo cervical/ 87
Ciclo vaginal/89
Ciclo vestibular/ 90
Variaciones en el pH/91
Referencias bibliogrficas/ 91
3. REGULACIN NEUROENDOCRINA DEL CICLO ESTRAL/ 95
Conceptos bsicos/ 95
Mecanismo de accin de las hormonas proteicas/100
Mecanismo de accin de las hormonas esferoides/101
Transporte de las hormonas/101
Hipotlamo/103
Factores de liberacin/103
Neuromediadores en el hipotlamo/107
Hipfisis anterior y su neurorregulacin/107
Gonadotropinas hipofisarias/108
Hipfisis posterior y su neurorregulacin/110
Funciones endocrinas del ovario/112
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
Progesterona/115
Relaxina/117
Oxitocina/117
lnhibinas/118
Andrgenos/119
Mecanismo feedback/120
Feedback de los esferoides gonadales sobre el hipotlamo (feedback largo)/121
Feedback hipofisario (feedback corto)/121
Feedback hipotalmico (feedback ultracorto)/121
Factores fisiolgicos que controlan el cuerpo lteo/121
Actividad lteo ltica del tero/123
Mecanismo de la lutelisis/129
Mecanismo de regulacin neurohormonal del ciclo estral/130
Gonadotropinas extrahipotisarias/130
Gonadotropina srica PMSG o gonadotropina corinica equina (eCG)/131
Gonadotropina corinica humana (hCG)/133
Propiedades antignicas de las gonadotropinas/133
Feromonas sexuales/133
Reconocimiento de la gestacin por la madre/134
Cuerpo lteo de gestacin/135
4. FECUNDACIN Y GESTACIN/142
Viabilidad de las clulas germinales/142
Sustancias inhibidoras de la motilidad de los espermatozoides/143
Transporte del esperma en el aparato genital femenino/143
Fenmeno de la capacitacin/145
Reaccin acrosmica/146
Cambios caractersticos de la motilidad durante la capacitacin/146
Fertilizacin/149
Clivaje/150
Migracin del huevo/151
Nutricin hemotrfica/153
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
Membranas fetales/153
Cordn umbilical/156
Placenta y placentacin/156
Clasificacin de las placentas/156
Fusin de las placentas en gestaciones mltiples/160
Circulacin sangunea fetal/160
tero grvido/164
Posicin y movimientos del feto durante la gestacin/165
Nmero de fetos en el tero/165
La madre durante la gestacin/168
Celo en la preez/168
Metabolismo/168
Respiracin y circulacin/169
Alimentacin durante la gestacin/169
Diagnstico de la gestacin/170
Mtodos de exploracin externa/170
Mtodos de exploracin interna/172
Caracterstica de la gestacin por meses/181
Posibilidades de error diagnstico/185
Diagnstico de la gestacin en la yegua/185
Migracin del conceptas/186
Otros mtodos de diagnstico/187
Ultrasonografa/187
Mtodos hormonales/187
Referencias bibliogrficas/189
5. EL PARTO/193
Mecanismo del parto/193
Signos de la proximidad del parto/ 201
Etapas del parto/ 203
Etapa de dilatacin cervical/ 203
Nomenclatura obsttrica/ 218
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
Presentacin/218
Posicin/219
Actitud/219
Transcurso del parto/ 220
Examen diagnstico de la parturienta/ 221
Nacimiento del feto/ 223
Inicio de la respiracin pulmonar/223
Estado cido-bsico del ternero recin nacido/ 225
ndice clnico del recin nacido/ 226
Ruptura del cordn umbilical/ 227
Tratamiento del cordn umbilical/ 227
Suministro de calostro/ 228
Atencin a la vaca recin parida/ 230
Traumas del canal del parto/ 230
Hemorragias/231
Otras medidas inmediatas con la madre/ 232
Puerperio/ 232
Reinicio de la actividad sexual posparto/ 235
Factores que influyen en el comienzo de la ciclicidad estral/ 236
Induccin del parto en la vaca/ 237
Introduccin del parto con prostaglandina/ 238
Introduccin y sincronizacin de partos con lidocana y prostaglandina/ 240
Parto de la yegua/ 241
Parto de la oveja/ 242
Parto de la cerda/ 243
Expulsin de las membranas fetales/244
Cras muertas/245
Asistencia al parto/245
Parto de la perra/ 245
Signos de la proximidad del parto/ 245
Primera fase del parto/ 246
Segunda fase del parto/ 246
Tercera fase del parto/247
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
Asistencia al parto/247
Parto de la gata/ 248
Referencias bibliogrficas/ 248
6. BIOTECNOLOGA DE LA REPRODUCCIN/254
Transferencia de embriones/ 254
Seleccin de las donantes/255
Seleccin de las receptoras/256
Tcnica de transferencia de embriones/ 257
Superovulacin de la donante/ 257
Sincronizacin del celo de la donante y de la receptora/ 262
Inseminacin de la donante/ 264
Bsqueda de los embriones/ 271
Evaluacin de los embriones/ 275
Implantacin de embriones/ 277
Aspectos epizootiolgicos de la transferencia de embriones/ 278
Conservacin de embriones/ 279
Esquemas de seleccin gentica MOET/282
Fecundacin in vitral 284
Clonaje/ 289
Tecnologa de la transferencia nuclear/ 290
Transgnesis/ 292
Referencias bibliogrficas/ 294
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
1. BASES MORFOLGICAS DE LA REPRODUCCIN
El estudio de la reproduccin en los animales domsticos requiere un pleno conocimiento
de las estructuras plvicas. La cavidad pelviana constituye el rea de trabajo en las labores
ginecobsttricas, tanto en la exploracin de los rganos genitales, como en la atencin al
parto, por ello se debe tener en cuenta las relaciones dimensionales en la futura
reproductora y en la parturienta, adems, las relaciones pelvimtricas ya sean internas
como externas avalan en buena medida la aptitud reproductora-productora de la hembra.
Una pelvis amplia presupone un parto fcil y una buena curva de lactancia.
De igual manera resulta conveniente referirse a la forma, la situacin y las relaciones de los
rganos que conforman el aparato genital, conocimientos bsicos y fundamentales para la
ubicacin y la exploracin manual de estas estructuras, en la que la subjetividad del tacto
puede limitarse a un mnimo de error, cuando la exploracin se practica con pleno dominio
anatmico de las formas y de las relaciones de los rganos que contiene.
La pelvis como conducto
El conducto pelviano es de vital importancia en la reproduccin, ya que este cinturn
oseoligamentoso, que contina a la cavidad abdominal, aloja a los rganos genitourinarios
(salvo en la hembra gestante), se adapta a la mejor disposicin para la cpula, sirve de
proteccin a la gestacin e interviene activamente en el parto, puesto que es atravesado por
el feto en su proceso de salida del claustro materno.
La porcin sea est estructurada por dos huesos simtricos, los coxales, unidos en su
regin ventral por la snfisis isquiopubiana y en la dorsal est presente el hueso sacro, todo
ello conforma el cinturn pelviano. Cada coxal est integrado por los huesos ilion, isquion
y pubis, que convergen en la cavidad acetabular o cavidad articular del fmur.
Estas formaciones seas sufren una lenta osificacin por sinostosis en la banda
cartilaginosa de la snfisis, la que dura aproximadamente cinco aos. En la vaca comienza
por la parte isquitica, para terminar en la pubiana, contrariamente a lo que sucede en la
yegua, que empieza por la pubiana, para concluir en la isquitica. En las novillas la snfisis
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
pubiana es bastante saliente y forma una cresta que, en ocasiones, puede dificultar el parto.
Esta cresta va desapareciendo con el transcurso del tiempo.
En los roedores la hormona relaxina tiene una fuerte actividad sinfisioltica, lo que permite
que en el momento del parto se separen ambos coxales.
Las estructuras blandas que completan la cavidad pelviana estn formadas por el ligamento
sacrocitico, este muestra dos porciones denominadas sacrospinoso y sacrotuberoso. El
ligamento sacrospinoso se extiende desde el borde del sacro hasta la espina isquitica y el
sacrotuberoso desde la parte dorsal del sacro hasta la tuberosidad isquitica.
En el ganado bovino los ligamentos pelvianos, en su borde posterior, estn cubiertos solo
por la piel, lo que da la posibilidad de palparlos como cuerdas tensas, las que se relajan
completamente en las horas que preceden al parto, esto constituye un signo importante
como prdromo del mismo. No sucede as en la yegua por hallarse dichos ligamentos
cubiertos de grasa que impide su palpacin. En casos de quistes ovricos foliculares
tambin se relajan y en condiciones normales permanecen firmes y tensos.
El conducto pelviano est limitado dorsalmente por el hueso sacro, que en la vaca, por lo
general, se le fusiona la primera vrtebra coxgea y en algunas ocasiones tambin la
segunda; el sacro se une a los huesos iliacos mediante sus carillas articulares. Lateralmente
el conducto se ve limitado por estos huesos, las espinas isquiticas y los ligamentos de la
pelvis de cada lado y ventralmente por el pubis y parte de los isquion que conforman el
suelo de la pelvis: cncavo en la vaca, lo que indica una mala disposicin para el parto y
convexo en la yegua, cuya disposicin es ms favorable, con la snfisis a todo lo largo de
su centro.
La disposicin acanalada del suelo de la pelvis de la vaca, no obstante sirve para acomodar
la vejiga durante el parto; en su porcin anterior se encuentra el estrecho anterior o
circunferencia pelviana, muy significativo desde el punto de vista obsttrico, est limitado
por la lnea terminal o borde, conformado por el promontorio (articulacin lumbosacra), las
lneas iliopectneas y el borde anterior del pubis. Su forma vara en las distintas especies de
acuerdo con el tipo de pelvis que posean (dolicoplvicas, platiplvicas o mesatiplvicas).
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
En su porcin posterior el conducto pelviano presenta el estrecho posterior de la pelvis, de
menos trascendencia desde el punto de vista obsttrico que el estrecho anterior, sus lmites
son: la articulacin sacrocoxgea, las arcadas isquiticas y el borde posterior del ligamento
sacrocitico de cada lado.
El largo de la cavidad pelviana lo determina la conjugada diagonal, la que se encuentra
entre el promontorio y la parte caudal de la snfisis pelviana. Cuanto ms larga sea la
pelvis, tanto ms difcil ser el parto.
Eje pelviano
El eje pelviano o eje de conduccin se encuentra en la unin de los centros de todas las
verticales trazadas del techo al suelo de la pelvis y tiene gran importancia en el proceso del
parto. La conduccin del feto durante la fase expulsiva se efecta siguiendo la direccin
del eje pelviano: parablico en la yegua (Figura 1.1), lo que le permite un parto rpido y
con escasas complicaciones; recto inicialmente en la vaca se continua hacia abajo y
termina ascendiendo ligeramente (Figura 1-.2), originando un parto demorado y con
frecuentes complicaciones. El conocimiento de estos aspectos resultan bsicos en los
auxilios al parto.
Figura: 1.1. Eje pelviano de la yegua (BA) parablico. Dimetro (CD) sacropubiano (Conjugada verdadera).
Dimetro (EF) sacroisquitico
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
Figura: 1.2. Eje pelviano de la vaca (BA) sigue una direccin en S con ascenso posterior. Dimetro (CD)
sacropubiano (Conjugada verdadera). Dimetro (EF) sacroisquitico
Diafragma plvico
El estrecho posterior de la pelvis est cerrado por una pared tendinosa, cncava hacia la
cavidad abdominal, que Benesch [1] denomin diafragma plvico. Est constituido por las
fascias plvica, perineal superficial y perineal profunda y los msculos constrictores
anteriores y constructor posterior de la vulva. Este diafragma divide el espacio plvico en
una porcin intraperitoneal y otra retroperitoneal.
El aparato genital a nivel de la unin del vestbulo con la vagina atraviesa el mencionado
diafragma que le forma un anillo, el anillo himenial, compuesto por el constrictor posterior
de la vulva (msculo bulbocavernoso) reforzado por haces musculares que se separan
directamente del elevador del ano y forman un msculo anular fusiforme del grosor de un
dedo pulgar.
Durante la cpula, este msculo estrecha la entrada de la vulva y comprime al pene, y
como est fijo al cltoris al contraerse eleva a este rgano, haciendo as los tactos ms
sensibles. A este tambin se debe el que la vulva se entreabra despus de la expulsin de la
orina, hacindose el cltoris externo.
El anillo himenial de fibras musculares estriadas se dilata durante el parto, de forma
mecnica, por las presiones que ejerce el feto a su paso por el canal del parto. Cuando no
se dilata adecuadamente, en especial en los partos con presentacin posterior, influye de
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
manera desfavorable en el desarrollo de este. Su deficiente involucin y retorno al tono
normal, en las horas que le suceden, predispone al prolapso del tero.
Dimetros del estrecho anterior de la pelvis
Por la naturaleza sea, del estrecho anterior o circunferencia craneal de la pelvis, sus
dimensiones casi no son susceptibles de aumentar durante el parto. De ah la necesidad de
conocer sus dimetros:
1. La conjugada verdadera, representada por la distancia entre el promontorio y el borde
anterior de la snfisis pubiana, la que determina la altura del estrecho anterior.
2. Los dimetros transversales o bisiliacos, medio, superior e inferior determinan el ancho
del estrecho anterior. El medio se encuentra entre las tuberosidades psodicas, el
superior va desde un ilion al otro limitando el tercio superior de la circunferencia y el
inferior que define el cuarto inferior
3. Los dimetros oblicuos o iliosacros, que van desde el extremo anterior de la
articulacin iliosacra de un lado, hasta la cresta iliopectnea del lado opuesto y se
cruzan en "X" en el centro del estrecho anterior. Estos dimetros tienen gran
importancia en la vaca por ser en esta especie mayor que los dimetros transversales,
por lo que constituyen va obligada del cinturn pelviano del feto, en el momento del
parto (Figura 1.3).
FIGURA 1.3 Estrecho anterior de la pelvis de la vaca con sus dimetros: conjugada verdadera (ab) Dimetro
bisiliaco superior (cd). Dimetros oblicuos o iliosacro (hf y ei).
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
Pelvis de las distintas especies domsticas
Teniendo en cuenta las relaciones de la altura (conjugada verdadera) con el ancho
(dimetros transversales) del estrecho anterior, las pelvis de las distintas especies
domsticas se han agrupado en tres tipos, que presentan mayor o menor facilidad o
disposicin para el parto:
Dolicoplvica
Este tipo de pelvis es propia de la vaca, la oveja, la cabra, la cerda y las siertas de razas
grandes. Presenta una circunferencia anterior alta y estrecha al superar la conjugada
verdadera al dimetro transversal medio, confirindole una forma oval.
En la cabra y la oveja debido a que las espinas y las tuberosidades citicas no son tan
prominentes, el techo de la pelvis es casi plano y el suelo es bastante corto y parejo, por lo
que la lnea de conduccin resulta ligeramente convexa, siendo mucho ms favorable para
el parto.
En la cerda con una pelvis similar a la de la vaca, con espinas y tuberosidades isquiticas
prominentes y suelo largo, a pesar de esa mala conformacin tocolgica, el parto se realiza
con facilidad gracias a la relativa pequeez de las cras y a que en los animales jvenes las
dos ltimas vrtebras sacras se articulan entre s con gran movilidad, adems, durante el
parto, las superficies articulares del sacro e ilion tienen amplios movimientos, y el feto, una
vez que ha atravesado el estrecho anterior, se encuentra en un espacio ms dilatable.
La perra tambin presenta una buena disposicin para el parto, ya que tiene el conducto del
parto y el hueso sacro, muy cortos, poca masa sea lateral y snfisis casi recta en un suelo
que se ensancha lateralmente hacia ambos isquion.
Mesatiplvica
La pelvis Mesatiplvica aparece en la yegua, las perras de presa y ratoneras. Con una gran
circunferencia anterior casi circular, tan ancha como alta, y escasas posibilidades de
trastornos durante el parto.
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
La pelvis de la yegua (Figura 1.5) con apfisis transversales del sacro poco prominente,
techo de la pelvis plano y suelo convexo y de poca longitud, que establece un eje de
conduccin parablico, se presenta como modelo en cuanto a amplitud y facilidades para el
parto.
Figura: 1.5. Pelvis mesatiplvica de la yegua. El dimetro transversal medio es similar a la conjugada
verdadera, lo que le da forma redondeada al estrecho, obsrvese en la vista lateral la poca longitud de la
pelvis y la poca altura de la cresta citica.
Distintos tipos de tero
Todos los vertebrados que ocupan en la escala zoolgica un lugar inferior al de los
marsupiales conservan conductos de Mller separados, los cuales se abren en la cloaca
definitiva. En los mamferos placentarios, por el contrario, estos conductos se fusionan en
grado variable, y con ello dan lugar a distintos tipos de tero.
Es de recordar que los conductos de Mller participan del desplazamiento que sufren las
crestas urogenitales y como consecuencia de esto cada conducto forma en su curso dos
incurvaciones que aproximadamente determinan tres regiones de diferentes potencialidades
futuras:
1. Una zona craneal (trompas).
2. Una zona media (fondo y cuerpo del tero).
3. Una zona caudal (cuello uterino y vagina posterior).
Tambin se debe recordar que el vestbulo vaginal se desarrolla del seno urogenital.
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
Segn el sitio donde se fusionen los conductos de Mller se originan los distintos tipos de
tero:
tero simple
Las regiones media y caudal de los conductos de Mller se fusionan y determinan un tero
piriforme, casi sin cuernos, estos rudimentarios reciben el nombre de trompas y toman
origen en la regin craneal de los conductos de Mller. Este tipo de tero es propio de la
mujer y de los primates superiores (Figura 1.7).
Figura: 1.8. tero bicornual subsepto de los rumiantes [7]
Figura: 1.9. tero bicornual no subsepto de la yegua [7]
tero dividido o bipartido
Este tipo de tero se presenta en la cerda y los carnvoros, en este la fusin de los
conductos de Mller se produce ms caudalmente, esto da lugar a cuernos muy largos y un
cuerpo uterino muy pequeo (Figura 1.10).
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
Figura: 1.1. tero dividido o bipartido de la cerda y los carnvoros.
tero doble
El tero doble es propio de los roedores y los lepridos, solamente estn fusionados los
conductos en su regin caudal, por lo que presentan dos cuernos uterinos completamente
separados, sin cuerpo, y con dos cuellos que desembocan separadamente en una vagina
comn (Figura 1.11).
FIGURA 1.11 tero doble de los roedores y los lepridos [7].
Los didelfos tienen dos vaginas que desembocan al exterior por separado, no existiendo
conexin entre los conductos (de aqu el nombre de didelfo). Esta disposicin se encuentra
en los marsupiales, como las zarigeyas. Adaptndose a esta peculiaridad de la hembra, el
macho tiene un pene bfido, capaz de insertarse de forma simultnea en las dos vaginas, lo
que ha dado origen a la conocida supersticin de que en las zarigeyas la copulacin se
lleva a cabo por la nariz [8].
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
Variaciones de la anatoma de los rganos genitales
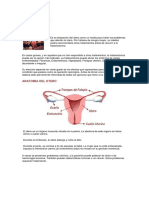
rganos genitales de la vaca
OVARIOS
Los ovarios miden aproximadamente de 3-4 cm. de longitud, de 2-3 cm. de ancho y de 1-2
cm. de espesor, con forma ovoide, que vana con la existencia u no de folculos y cuerpos
lteos y la posicin que estos tengan en el ovario. La presencia de estas formaciones le da a
los ovarios cierta forma redondeada.
En las vacas estos son relativamente pequeos en relacin con los de otras especies
domsticas; en la novilla tienen menor tamao y en la ternera recin nacida el ovario
izquierdo es en general algo ms grande que el derecho, mientras que en la hembra madura
el ovario derecho es con frecuencia el mayor.
Los ovarios estn situados en la hembra vaca a nivel de la bifurcacin de los cuernos,
Junto a su pared lateral externa, a unos 30-45 cm. de la abertura vulvar. Se hallan unidos a
los cuernos por el mesovario, que es la porcin anterior del ligamento ancho. El ligamento
ovrico une la extremidad uterina del ovario con la del cuerno o pice.
La posicin de los ovarios es muy inconstante dado su modo de insercin.
Cada ovario o ambos, pueden desviarse transversalmente en todas direcciones, hasta el
lmite mximo permitido por el mesovario.
El ovario tiene una corteza, zona cortical, en la que estn situados los folculos y una zona
medular, que contiene los vasos y los nervios. La mayor parte de la superficie de la
glndula est cubierta de epitelio germinativo, el epitelio peritoneal se encuentra limitado a
una zona estrecha a lo largo del borde de insercin, lo que permite que la ovulacin se
produzca en cualquier lugar de la superficie ovrica.
OVIDUCTOS
Los oviductos son un par de tubos flexuosos de 20 - 30 cm. de longitud que se extienden
desde las extremidades de los cuernos hasta los ovarios. Estn encargados de la captacin
del vulo y de su conduccin hasta el tero. Sin embargo, no se hallan en comunicacin
directa con los ovarios sino ms bien en parte contiguos a ellos y en parte insertos en estos.
Su orificio uterino es bastante grande e infundibuliforme.
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
Los oviductos, trompas uterinas o trompas de Falopio, se dividen en tres segmentos:
1. Pabelln, extremo ovrico del oviducto constituido por la reunin de fimbrias, que se
ensancha formando un embudo, est encargado de captar el vulo, puede llegar a tener
de 5-7 mm de dimetro.
2. Ampolla tubrica, lugar donde se produce la fecundacin, tiene un dimetro de 3-5
mm.
3. Istmo, el ms largo y estrecho de los tres segmentos (0,8-1 mm de dimetro), se
extiende desde la ampolla al cuerno correspondiente.
Los oviductos estn sostenidos por un pliegue peritoneal derivado de la parte lateral del
ligamento ancho. Las fimbrias se hallan fijadas al borde libre de este ligamento
(mesosalpinx), lo que facilita que cuando se produzca la ovulacin el pabelln con
movimientos activos se aproxime al ovario y posibilite de este modo la captacin del
vulo. Dado el origen (conductos de Mller) y estructura, similares al tero, los oviductos
o trompas pueden ser considerados como prolongaciones de este.
TERO
El tero de la vaca, como el de la mayora de los animales domsticos, es un rgano
bicorne (Figura 1.12). En el animal no gestante, est situado normalmente a unos 25-40
cm. por delante de la abertura vulvar en posicin anterior con respecto al cuello uterino. El
cuerpo del tero, que puede tener de 9-12 cm. de dimetro transversal, por lo general, tiene
nada ms que 2-5 cm. de longitud, aunque a la palpacin da la sensacin de ser mayor
debido al ligamento intercornual muy desarrollado, que prolonga virtualmente la longitud
del cuerpo uterino hacia adelante.
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
Figura: 1.12. rganos genitales de la vaca [9]
La parte anterior del cuerpo uterino est dividida en dos cuernos que estn unidos en una
distancia de 10-15 cm., lo que contribuye con el ligamento intercornual a que el cuerpo del
tero parezca ms largo de lo que realmente es. A nivel del punto de separacin de los
cuernos, estos tienen un dimetro de 3-4 cm. y se adelgazan e incurran poco a poco,
primero hacia fuera y abajo, luego hacia atrs y arriba, para continuarse con los oviductos.
Las extremidades de los cuernos son agudas por lo que su unin con las trompas en la
conjuncin tero-tuba rica no es abrupta como en la yegua. El orificio tubrico de las
trompas es bastante grande e infundibuliforme. Estos tienen desde el punto de separacin
cornual de 20-35 cm de longitud, la que vara con la edad del animal, su talla, el nmero de
gestaciones y otros factores.
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
En la novilla no gestante el tero generalmente se encuentra en la cavidad pelviana y solo
una parte de los cuernos se extiende ms all de la entrada de la pelvis. En las vacas viejas,
gordas, de razas lecheras o que hayan tenido varios partos, el tero tiende a colgar en la
cavidad abdominal, y puede aparecer casi totalmente en ella en vacas que han tenido varios
partos,
CUELLO UTERINO
Et cuello uterino o crvix es un esfnter muscular situado entre el tero y la vagina, de
forma cilndrica y paredes gruesas, de 8-10 cm de longitud y de 1,5-2 cm de ancho en las
novillas, aumentando de longitud unos 10-15 cm y de ancho 3-5 cm en las vacas, en
funcin de la edad y el nmero de partos, la forma que adquiere es piramidal de base
posterior; en la vaca resulta el punto de referencia ms significativo para la localizacin del
aparato genital durante su exploracin por va rectal.
Las vacas Ceb y sus mestizas presentan mayor grosor del cuello por hipertrofia del tejido
colgeno. En ocasiones este toma dimensiones considerables, como el caso de una vaca
Santa Gertrudis en la que el cuello tena aproximadamente de 25-30 cm de longitud y de
12-15 cm de ancho. Lo ms notable de estos casos es que son fecundadas y paren sin
dificultad aun en casos extremos como el citado.
El conducto cervical muestra prominencias transversas entrelazadas llamadas anillos, muy
prominentes en las vacas. En su abertura vaginal, en la flor radiada, dicho conducto tiene
el primer anillo, que se denomina anillo de Burdi, formado por tres repliegues o labios
carnosos (el superior o transversal y dos laterales de direccin oblicua) que tienen gran
importancia en el diagnstico de la cervicitis.
VAGINA
La vagina se extiende desde el cuello del tero hasta el vestbulo a travs de la cavidad
pelviana. En el animal no gestante la vagina y el vestbulo vaginal juntos miden de 15-30
cm. Se relaciona por la parte dorsal con el recto, por la ventral con la vejiga y la uretra y
por la lateral con la pared pelviana. Durante la gestacin el tero desciende en la cavidad
abdominal, tirando de la vagina que puede duplicar su longitud.
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
La pared vaginal es delgada, pero muy resistente y elstica, por lo que durante el parto, la
dilatacin de la vagina solo queda limitada por las paredes de la cavidad pelviana.
En la pared ventral de la vagina, entre las tnicas muscular y mucosa, se encuentran
generalmente los conductos de Gartner, vestigios de los conductos de Wolff. Tienen inters
clnico porque con alguna frecuencia pueden dar lugar a quistes.
VULVA
Se designa como vulva a la parte posterior del aparato genital femenino; comprende el
vestbulo vaginal y los labios vulvares. Por dentro de la comisura inferior de los labios
vulvares se halla una pequea formacin, el cltoris, homlogo embriolgico del pene, el
cual est formado por dos pequeos cuerpos cavernosos erctiles que se unen (cuerpo) y
terminan en un glande rudimentario. El cltoris en su totalidad tiene unos 10 cm de
longitud, pero solamente es visible la extremidad de este rgano. Durante el acto sexual, el
cltoris es capaz de cierto grado de ereccin, sin presentar nunca la turgencia y la
movilidad que alcanza en la yegua.
El orificio uretral externo, se halla de 10-12 cm de la comisura ventral, a la altura del
himen. Debajo de este existe un saco ciego, el divertculo suburetral, que mide cerca de 3,5
cm. de longitud y puede permitir fcilmente la introduccin de un dedo. La forma y la
posicin de este divertculo hay que tenerla en cuenta cuando se vaya a practicar el
cateterismo vesical en la vaca, ya que si se pasa el catter a lo largo de la pared ventral de
la vulva como se hace en la yegua, penetra siempre en el citado divertculo en lugar de
hacerlo en la uretra.
A cada lado del vestbulo, por fuera y ligeramente por detrs del orificio uretral se
encuentran las desembocaduras de las glndulas vestibulares mayores o glndulas de
Bartholin, al parecer de poca importancia en la vaca.
La vulva tiene labios gruesos y arrugados con una comisura ventral que termina en un
borde agudo, proyectado hacia afuera, en si que existe un pequeo mechn de pelos y la
comisura dorsal es redondeada.
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
IRRIGACIN SANGUNEA
El aparato genital est irrigado por las arterias ovrica, uterina y urogenital.
Arteria ovrica. Antigua arteria tero ovrica, es pequea y nace de la cara ventral de la
aorta. Corre por la parte anterior del ligamento ancho del tero y va a irrigar al ovario, da
una rama uterina que se dirige al borde cncavo del cuerno uterino, al que irriga,
anastomosndose con la arteria uterina.
Arteria uterina. Muy voluminosa nace de la iliaca interna, de un tronco comn con la
umbilical, desciende sobre la pared lateral de la pelvis un corto trecho por detrs de la
iliaca externa y avanza por el ligamento ancho del tero hasta alcanzar la cara dorso lateral
de este, inmediatamente por delante del cuerpo. Se distribuye principalmente por el cuerpo
del tero.
Arteria urogenital. Antigua arteria uterina posterior, nace de la pudenda interna, irriga la
parte posterior del tero y emite ramas a la vagina y a la vejiga y termina constituyendo la
arteria del cltoris [10].
ELEMENTOS DE SUJECIN
De la porcin dorsal de la cavidad pelviana parten los ligamentos anchos (mesometrio) y se
insertan en la curvatura menor de los cuernos en su borde latero inferior, con lo que estos
quedan ms apoyados que suspendidos y hasta cierto punto en un equilibrio inestable
(fenmeno que explica que la torsin uterina sea ms frecuente en la vaca).
La arteria uterina que recorre el espesor de los citados ligamentos, procede de la iliaca
interna y tienen importancia en el diagnstico de la gestacin.
rganos genitales de la yegua
OVARIOS
Los ovarios de la yegua tienen forma arrionada a causa de la presencia de la fosa de
ovulacin y su consistencia elstica firme. El peso entre 40-80 g con 7-8 cm de longitud y
3-4 cm de ancho. Et borde de insercin o mesovrico es convexo y el borde libre presenta
una depresin estrecha, la fosa de ovulacin. Estn situados en la regin sublumbar a nivel
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
de las cuarta y quinta vrtebras lumbares a un travs de mano por detrs de los riones y
separados del plano medio.
El ovario izquierdo est comnmente 2 o 3 cm ms atrs que el derecho, pero se halla casi
siempre ms cerca del rin correspondiente. Por lo general en la yegua que no est en
gestacin se hallan en contacto con la pared abdominal lumbar.
La estructura del ovario de la yegua es caracterstica, ya que no presenta una zona cortical
en la que estn presentes los folculos; estos se distribuyen por el interior de la glndula y
deben hacer el recorrido momentos antes de la ovulacin, hasta la fosa de ovulacin, nico
lugar donde pueden hacer eclosin, debido al espesor y consistencia de la capa albugnea
que rodea el resto del ovario.
Otra caracterstica del ovario de la yegua es que el cuerpo lteo no forma relieve en la
superficie del ovario como sucede en la vaca y en la cerda, sino que est situado en el
interior de la glndula.
A diferencia de la vaca, en la yegua los ovarios constituyen, durante la exploracin rectal,
el punto de referencia ms importante para la localizacin del aparato genital.
OVIDUCTOS
Los oviductos tienen una longitud de 25-30 cm y en su trayecto experimentan un notable
serpenteo. Cada uno est envuelto en un pliegue peritoneal, derivado de la cara externa del
ligamento ancho, denominado el mesosalpinx, el que cubre en gran parte el lado externo
del ovario y forma con el ligamento ancho una bolsa, llamada bolsa ovrica.
En el mesosalpinx se hallan tbulos flexuosos ciegos, que constituyen el paroophoron,
vestigio del conducto de Wolff. Estos son ms evidentes en las yeguas adultas jvenes y
tienden a desaparecer con la edad, no es raro que den lugar a quistes.
El pabelln del oviducto de la yegua se caracteriza por los amplios pliegues que presenta
en la fase de reposo sexual y se adapta perfectamente sobre la fosa de ovulacin, por lo que
solo, causas mecnicas, tumores, quistes, etc., pueden determinar la cada del vulo en la
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
cavidad abdominal. En sus bordes es frecuente encontrar quistes pedunculados y las
hidtides de Morgagni.
TERO
El tero de la yegua es bicorne, no subsepto (Figura 1.13), es decir, no tabicado; ambos
cuernos aparecen unidos en su base por el ligamento intercornual, que no es tan
desarrollado como en la vaca. Estn situados enteramente en el abdomen y comprimidos
contra los msculos sublumbares por los intestinos (ciego, porcin izquierda del colon
mayor, el colon menor y el intestino delgado). El borde dorsal es algo cncavo y est unido
a la regin sublumbar por el ligamento ancho; el borde ventral es convexo y libre.
El cuerpo del tero est situado parte en la cavidad abdominal y parte en la pelviana. Su
longitud es de 18-20 cm y su dimetro de unos 10 cm. Su cara dorsal se relaciona con el
recto y otras porciones del intestino; la ventral est en contacto con la vejiga y tiene
relaciones inconstantes con varios segmentos del intestino.
La posicin del cuerpo del tero es variable, especialmente en lo referente a su porcin
anterior. A menudo est comprimido contra el recto y puede ser desviado hacia ambos
lados, ms frecuentemente hacia el izquierdo, por la flexura pelviana del colon mayor o por
asas del colon menor.
FIGURA 1.13 rganos genitales de la yegua [11]
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
CUELLO UTERINO
El cuello uterino de la yegua es un pequeo conducto de 4-7 cm de largo y de 3,5-4,5 cm
de ancho, de forma cilndrica algo aplanada, su abertura posterior se proyecta en la cavidad
vaginal, en la que termina y forma una serie de repliegues, la flor radiada, delimitados
circularmente del fondo de la vagina por un profundo surco, el fornix vaginalis. Su
posicin es paralela al eje de la pelvis.
El conducto cervical es permeable para el dedo, el que lo puede ensanchar bastante sin
dificultad. En la lnea meda puede comprobarse con frecuencia un pliegue de la mucosa,
que desde la pared superior de la bveda vaginal se dirige a la portio, a veces dicho pliegue
solo est insinuado. Desde la parte interior del orificio uterino existe un rafe central de
mucosa que se bifurca en la direccin del suelo de la vagina.
VAGINA
La vagina alcanza de 15-25 cm de longitud y unos 12 cm de dimetro, de paredes gruesas
pero muy dilatables. Se relaciona dorsalmente con el recto, ventralmente con la vejiga y la
uretra y lateralmente con la pared pelviana.
Externamente el tercio anterior de la vagina est cubierto por el peritoneo los dos tercios
posteriores por un tejido conjuntivo laxo, un plexo venoso una cantidad variable de grasa,
de ese modo el peritoneo forma por la regin dorsal un saco rectovaginal entre la vagina y
el recto y por la ventral uno vesicovaginal entre la vagina y la vejiga, quedando la mayor
parte de la vagina retroperitoneal y casi completamente retroperitoneal cuando el recto y la
vejiga se encuentran llenos.
VULVA
La vulva o seno urogenital (vestbulo vaginal) mide de 10-12 cm de longitud desde el
orificio uretral externo hasta la comisura ventral, por el dorso es mucho ms corta. Se
relaciona dorsalmente con el recto y el ano ventralmente con el suelo de la pelvis y
lateralmente con el ligamento sacrocitico, el msculo semimembranoso y la arteria
pudenda externa.
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
La hendidura vulvar de 10-12 cm de altura, compuesta por dos labios redondeados
prominentes, los labios vulvares, los que se unen arriba en ngulo agudo y forman la
comisura dorsal, y abajo la gruesa y redondeada comisura ventral.
Los labios estn cubiertos por la parte exterior de piel fina, lisa, pigmentada, desprovista de
pelos y con abundantes glndulas sudorparas y sebceas, por lo que es untuosa al tacto;
por la interior con una delgada membrana mucosa desprovista de glndulas. Cuando se
separan los labios se ve el glande del cltoris, cuerpo redondeado de aproximadamente 2,5
cm de ancho que ocupa la llamada fosa del cltoris. Est cubierto por un delgado
tegumento pigmentado, con el que reviste la fosa, este constituye el prepucio. E cuerpo de
tejido erctil semejante al del cuerpo cavernoso del pene, tiene una longitud de unos 5 cm.
y su dimetro es aproximadamente como el de dedo meique.
El msculo constrictor anterior de la vulva se inserta en la base del cltoris y tiene forma de
esfnter. Durante la cpula comprime los labios vulvares, eleva el cltoris y aumenta de ese
modo el contacto entre los genitales.
El constrictor posterior tiene como misin estrechar la cavidad vulvar, al mismo tiempo
que el vestbulo uretral. En la extremidad anterior de la pared ventral de la vulva a 10 o 12
cm de la comisura ventral, se halla el orificio uretral externo, que admite fcilmente un
dedo y es muy dilatable.
rganos genitales de los pequeos rumiantes
OVARIOS
Los ovarios de los pequeos rumiantes son proporcionalmente mayores que los de los
grandes rumiantes, tienen forma de almendra y su longitud es de 1,5 cm aproximadamente,
estos permanecen muy prximos al extremo craneal de los cuernos, a la altura de la cresta
del pubis y presentan una bolsa ovrica, ancha y aplanada.
OVIDUCTOS
Las trompas uterinas u oviductos miden de 10-15 cm de longitud, pasan gradualmente a la
porcin ms estrecha de los cuernos y entre estos no existe demarcacin.
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
TERO
El tero es tambin bicorne y subsepto como el de la vaca, los cuernos de 10-12 cm de
longitud, forman una espiral cerrada y sus porciones posteriores se unen en 3 cm o ms de
extensin. Las carnculas mucho menores que las de la vaca, poseen una depresin en su
cara libre y suelen estar pigmentadas.
El cuerpo del tero tiene aproximadamente de 1,5 - 2 cm de longitud. El cuello mide cerca
de 4 cm de longitud; la luz del conducto cervical est cerrada por prominencias y
depresiones recprocas de la mucosa.
El orificio externo se halla en la parte ventral de la vagina y no en su centro y fondo y est
recubierto por la plica cervicalis, carnosa y prominente, sobre todo en la oveja.
VAGINA
La vagina mide unos 8 cm de longitud y contiene numerosos folculos linfticos en su
porcin ventral. En ella desemboca la uretra, corta y provista de un pequeo divertculo
suburetral.
VULVA
La vulva mide de 2,5-3 cm de longitud. Los labios, arrugados, estn estira- dos en la
comisura ventral, en forma de pico de jarra, provisto de pelos. El cltoris es corto y dbil,
semejante al de la vaca.
rganos genitales de la cerda
OVARIOS
Los ovarios son redondeados y adquieren forma de mora, por el gran nmero de folculos
que maduran a la vez. Estn situados como en los rumiantes en el estrecho anterior de la
pelvis o prximos a ella, pero su posicin es muy variable pudiendo encontrarse cerca de
los riones (3-5 cm).
Los folculos maduros tienen un dimetro de 7-8 mm y los cuerpos lteos alcanzan de 1215 mm. La dehiscencia folicular se acompaa de abundante hemorragia. Debido a la gran
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
extensin del mesosalpinx los ovarios estn casi completamente rodeados por la bolsa
ovrica.
OVIDUCTOS
Las trompas de Falopio u oviductos tienen gran desarrollo (15-30 cm), son flexuosos, con
pabelln muy desarrollado que forma una especie de bolsa, con amplia abertura abdominal.
Su extremidad uterina se contina insensiblemente con la extremidad afilada del cuerno.
TERO
El tero es dividido o bipartido, el cuerpo es corto y el cuello relativamente largo (Figura
1.14). Los cuernos uterinos son largos y flexuosos, libremente mviles gracias a la longitud
de sus ligamentos; fuera del estado de gestacin estn dispuestos en numerosas asas,
similares a las del intestino delgado, con una longitud de 1,2-1,5 m, que puede alcanzar
ms de 2 m durante la gestacin. Las extremidades de los cuernos se adelgazan hasta
presentar el dimetro de las trompas.
FIGURA 1.14 rganos genitales de la cerda [11]
CUELLO UTERINO
El cuello uterino pasa a la vagina sin transicin, de modo que no forma una verdadera
porfo. El conducto cervical presenta pequeas prominencias redondeadas, algunas de las
cuales se ensamblan en forma de cremallera que cierra firmemente el tero.
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
VAGINA
La vagina de 12-15 cm de longitud es fuertemente musculosa, de pequeo calibre, su
mucosa posee tubrculos que adquieren turgencia durante el celo, estos en el momento del
coito se adhieren y enlazan a las espirales del pene, y as aumenta el contacto sexual. La
parte posterior de la uretra est fusionada con la vagina y produce una elevacin en el suelo
de esta.
VULVA
La vulva mide cerca de 7 cm de longitud y sus labios son gruesos. La comisura dorsal es
redondeada, pero la ventral se prolonga en forma de lengua. El cltoris muy largo, tino y
rojizo se proyecta sobre su fosa.
rganos genitales de la perra
OVARIOS
Los ovarios son pequeos y alargados de 1-2 cm de longitud; estn situados por detrs del
polo posterior de los riones o en contacto con estos, aproximadamente a la mitad de la
distancia entre la ltima costilla y el ngulo externo del ilion, por tanto a nivel de las
tercera y cuarta vrtebras lumbares. El ovario derecho se halla entre el duodeno y la pared
lateral del abdomen y el izquierdo est relacionado lateralmente con el bazo. Los ovarios
estn encerrados en un pliegue peritoneal, la bolsa ovrica, que presenta ventralmente una
abertura en forma de hendidura y sus paredes tienen un depsito de grasa.
OVIDUCTOS
Las trompas u oviductos tienen una longitud de 6-9 cm, ligeramente flexuosos, casi rodean
al ovario; su orificio abdominal es bastante grande y se abre en la bolsa ovrica, en cambio
el uterino es muy pequeo.
TERO
El tero tiene un cuerpo muy corto (2-3 cm) y los cuernos bastante largos y estrechos, que
se separan del cuerpo uterino en forma de "V", de 12-15 cm de longitud (Figura 1.15); son
rectos con la peculiaridad que el derecho es algo ms largo que el izquierdo y se
encuentran enteramente en la cavidad abdominal.
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
Cuando el tero est grvido los cuernos presentan dilataciones, las ampollas, que
contienen los fetos y estn separadas por constricciones. El tero grvido se halla sobre la
pared abdominal ventral y hacia el trmino de la gestacin se extiende hacia delante hasta
el estmago y el hgado, donde sufren lateroflexiones fisiolgicas, al igual que sucede en la
cerda.
FIGURA 1.15 rganos genitales de la perra
CUELLO UTERINO
El cuello uterino es de paredes gruesas, corto y tiene un conducto cervical estrecho. En la
porcin ventral del orificio externo del cuello existe una eminencia cilndrica de 5-8 mm de
longitud que se halla en una depresin de la pared vaginal.
VAGINA
La vagina es relativamente larga cuya membrana mucosa forma pliegues longitudinales.
En la pared dorsal hay un pliegue muy manifiesto que va a formar, en cierto modo, la parte
dorsal del orificio externo del cuello del tero.
VULVA
La vulva tiene labios gruesos y estos forman una comisura inferior aguda, dirigida hacia
atrs y provista, por lo general de pelos. El cuerpo del cltoris es ancho y plano. Su
estructura no es erctil sino que est infiltrada de grasa. El glande del cltoris est
compuesto tambin de tejido erctil y est situado en una gran fosa, la fosa clitoroidea.
Las dimensiones medias de los rganos genitales de todas estas especies aparecen en la
tabla 1.1.
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
TABLA 1.1 DIMENSIONES MEDIAS DE LOS RGANOS GENITALES EN
ALGUNAS HEMBRAS DOMSTICAS (cm)
Especie
Ovarios
Vaca
L: 3 - 4
A: 2-5
Cuernos
Cuerpo
Cuello
Vagina
35
2-5
8-10
15-30
25
18-28
3-7,5
15-20
E: 1,5-2
Yegua
L5-8
E:2,5-4
Oveja
L1-5
10-15
1.5-2
Cerda
L2-3
1,2-1,5
10
10-12
Pena
L: 2
12-15
2-3
L: largo
A: alto
E: espesor
Situacin intravital de los rganos genitales
El conocimiento de la situacin intravital de los rganos genitales de la vaca y de la yegua,
es un elemento bsico e indispensable para realizar el examen rectal.
La posibilidad que brindan estas especies, de poder palpar minuciosamente sus rganos por
va rectal es sin duda una extraordinaria ventaja de que dispone el Mdico Veterinario. Esto
constituye un mtodo prctico para e diagnstico certero de la gestacin, de la actividad
ovrica y de mltiples afecciones del aparato genital. Representa, por tanto, un valioso
resorte en su lucha contra la infertilidad y la subfertilidad.
Situacin intravital de los rganos genitales de la vaca
El tero vaco de la vaca sana, cuyos cuernos estn arroyados hacia abajo y afuera como
cuernos de carnero, generalmente est situado sobre el borde de pubis, aunque algunas
porciones de este se pueden situar en la cavidad abdominal; se relaciona con el recto por la
regin dorsal (Figura 1.16).
En la mayor parte de las vacas Guernsey y en muchas Shorthorn, el rgano es
predominantemente abdominal [11]. Tambin es abdominal en vacas viejas y gordas. En
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
las novillas y en las vacas subalimentadas se encuentra situado en su totalidad en la
cavidad pelviana.
El tero usualmente es dorsal o lateral y craneal a la vejiga urinaria, por lo que cuando esta
se encuentra llena lo puede desplazar.
Durante la gestacin el tero aumenta gradualmente de tamao y peso, y avanza y cae al
suelo de la cavidad abdominal. El primer cambio de posicin que se observa es un
desplazamiento con frecuencia hacia un lado, de la gran curvatura de los cuernos. A esto
sigue un descenso hacia la cavidad abdominal que se inicia alrededor de los 75 das de
gestacin y se completa aproximadamente entre los das 130 y 140. El ascenso por
crecimiento de feto comienza a los 7 o 7 1/2 meses.
La vagina est ubicada en la cavidad pelviana, dorsalmente se relaciona con el recto y
ventralmente con la vejiga urinaria.
Los ovarios en la vaca vaca se sitan laterocaudalmente a la curvatura mayor de los
cuernos, a linos 10 cm de distancia de la lnea media y de 2-5 cm por delante de la espina
iliaca.
FIGURA 1.16 Situacin intravital de los rganos genitales de la vaca [9].
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
Al aumentar el peso del tero este desciende y ejerce traccin sobre el ligamento ancho.
Como resultado los ovarios se desplazan anterior y medialmente, pero ante todo
ventralmente. El descenso de los ovarios los aleja de la persona que practique el examen
rectal hasta hacerlos inalcanzables.
Todo el rgano se halla fijo a la porcin dorsal de la cavidad pelviana por los ligamentos
anchos del tero (mesometrios). Dichos ligamentos se insertan ventrolateralmente en la
curvatura menor de los cuernos y lateralmente al cuerpo y al cuello uterino.
El borde anterior de los ligamentos es oval y forma el ligamento tero-ovrico y el
mesovario (ambos unidos forman la bolsa ovrica). Los ovarios estn suspendidos
aproximadamente 5 cm a los lados del extremo ovrico del cuerno uterino correspondiente.
Situacin intravital de los rganos genitales de la yegua
En la yegua vaca y al principio de la gestacin, los ovarios se encuentran en la regin
sublumbar 5-10 cm por delante de la espina iliaca, a un travs de mano por detrs de los
riones y separados del plano medio; se hallan unidos a los msculos lumbares por el
mesovario. La amplitud con que se pueden desplasar los ovarios est limitada por la
longitud del mesovario. La incapacidad de localizarlos en su posicin normal es muy
sugestiva de gestacin avanzada.
El tero de la yegua posee un cuerpo relativamente grande y cuernos pequeos que se unen
al cuerpo casi perpendicularmente, dando al rgano una forma de "T". Los cuernos son
convexos, con la curvatura mayor mirando hacia el frente, hacia abajo y lateralmente. Los
cuernos y el cuerpo estn aplanados por las partes dorsal y ventral. El borde anterior del
cuerpo y los cuernos se encuentran a nivel del reborde plvico, donde forma un arco casi
paralelo al compuesto por las espinas iliacas y el hueso pbico [11].
El tero en esta especie se observa suspendido, dorsal a la vejiga urinaria, por el ligamento
ancho, que est unido a su pared abdominal en la regin sublumbar (Figura 1.17).
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
Durante la gestacin avanzada los ovarios se desplazan hacia abajo, adelante y hacia la
lnea media en posicin fija. Los cuernos aparecen flexionados hacia un lado sobre la
superficie fluctuante que representa el tero.
FIGURA 1.17 Situacin intravital de los rganos genitales de la yegua [11]
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
REFERENCIAS BIBLIOGRFICAS
1). BENESCH, R: Obstetricia y ginecologa veterinarias, Editorial Labor, S.A., Madrid,
1963.
2). Rizo, J.M.; 0. FERNNDEZ; A. Gil y R. GUIROLA: Factores de alto riesgo
obsttrico en condiciones de manejo de Cuba, (indito).
3). RICE, LE. y J.N. WILTBANK: Factors affectin dystocia in beef heifers, Am. Vet.
Med. Assoc., 161(11):1348,1972.
4). GAINES, J. D.; D. PESCHEL; R.G. KAUFFMAN, et al.: Pelvic growth calf birth
weight and dystocia in Holstein x Hereford Leifers, Therogenology, 40 (1)33,1993.
5). VAN DONKERSGOED, J.; C.S. RIBBLE; C.W. BOOKER et al.: The predictive
valu of pelvimetry in beef cattle, Can. J. Vet. Res., 57(3):170,1993.
6). HINDSON, J.C.: Quantification of obstetric traction, The Veterinary Record,
102:327-332,1978.
7). NALBANDOV, AV.: Fisiologa de la reproduccin, Ed. Acribia, Zaragoza, 1969.
8). ZEMJANIS, R.: Reproduccin animal. Diagnstico y tcnicas teraputicas, Ed.
Limusa-Wiley, S.A., Mxico, 1966.
9). HOLY, L: Biologa de la reproduccin bovina, Ed. Pueblo y Educacin, La Habana,
1975.
10). GETTY, R.; S. SISSON y GROSSMAN: Anatoma de los animales domsticos, 5ta,
Ed., Salvat Editores S. A., Barcelona, 1984.
11). SISSON, S.: Anatoma de los animales domsticos, Salvat Editores, S.A., 1977.
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
2. FUNCIONES REPRODUCTIVAS
El estudio de las peculiaridades reproductivas de cada especie es una premisa bsica y
fundamental para enfrentar con amplias probabilidades de xito, la dinmica de la
actividad que los mtodos modernos de reproduccin requieren. La creciente demanda de
protenas de alto valor biolgico (leche, carne) constituye un verdadero acicate para
optimizar la explotacin al mximo de sus capacidades pro-creativas.
Si bien es cierto que se ha avanzado en el dominio y la adecuacin de la parte masculina,
no sucede lo mismo con la femenina, en la que la obtencin de una cra dentro de los
parmetros deseables (12-13 meses en la vaca) representa una meta difcil de obtener y
mucho ms de mantener; mirando retrospectivamente se puede constatar que en los ltimos
aos se ha avanzado y que existen grandes perspectivas en un futuro no muy lejano.
El conocimiento de los diversos factores que regulan la ciclicidad estral ha permitido la
aplicacin prctica de la progesterona y de eficaces gestgenos, la prostaglandina F; a y sus
valiosos anlogos, la gonadotropina srica, la FSH y la GnRH, y otras que abren un gran
campo a la utilizacin del potencial reproductivo de las hembras. No obstante, solo el
dominio de las caractersticas ms sutiles del ciclo estral, pondrn al hombre en
condiciones de lograr de cada hembra, lo que ya se obtiene en productividad reproductiva
de los machos.
Ciclo reproductivo
Desde el punto de vista de la reproduccin y, sobre todo, de la utilizacin econmica de la
hembra,
es
necesario
establecer
tres
etapas:
prerreproductiva,
reproductiva
posreproductiva.
En la misma medida en que se persigue el mayor rendimiento econmico de la ganadera
moderna, se le da mayor valor a las dos primeras etapas (prerreproductiva y reproductiva),
puesto que la cra intensiva implica eliminar las hembras cuando comienzan a declinar en
sus capacidades productivas y reproductivas.
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
Etapa pre-reproductiva
El perodo que antecede a la vida reproductiva de los animales domsticos,
especficamente, cuando estos se cran con fines econmicos, tiene especial importancia,
por requerir intensos cuidados, gastos de instalaciones y alimentos, as como para preservar
su salud, que no sern reintegrados hasta tanto entren en produccin. Adems, es el perodo
idneo para iniciar el proceso de seleccin, tan necesario en la ganadera moderna.
Los ovarios de la novilla prepber son activos y contienen folculos en crecimiento antes
de que muestren actividad estral. En general, estos folculos sufren regresin y atresia,
incluso hasta la pubertad continan olas de desarrollo y regresin folicular [1].
Las gnadas de animales prepberes son capaces de presentar respuesta superovulatorias,
frente a la estimulacin exgena de gonadotropina, aun cuando se administre mucho antes
de la edad normal de pubertad, lo que resulta de gran inters en la transferencia de
embriones, cuestin que permite ganar tiempo en el avance gentico. Ms an la glndula
hipfisis de animales recin nacidos y hasta la de los fetos responde a la estimulacin
exgena de GnRH, secretando y liberando gonadotropinas y como lo hara un animal
pospber [2].
Etapa reproductiva
La etapa reproductiva se inicia al establecerse la pubertad y los ciclos estrales. Durante esta
se producen las gestaciones y los partos, eventos indispensables para que se inicien los
ciclos productivos, por lo que sin dudas, constituye la fase ms importante, desde el punto
de vista econmico.
Etapa posreproductiva
Se caracteriza la etapa posreproductiva, por la declinacin de las funciones reproductivas
del animal, aunque en ellos no se manifiesta la menopausia con el cese completo de estas
funciones como sucede en la mujer. No obstante, en la medida en que el animal envejece,
disminuye su produccin de hormonas gonadales, con lo que pierde el efecto anablico de
las protenas, causa de la emaciacin de la edad avanzada [2].
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
Por ser la etapa posreproductiva, una fase negativa desde el punto de vista econmico, las
hembras de los animales domsticos, no son mantenidas hasta el climaterio, con la
excepcin de las de las especies afectivas.
La declinacin de las funciones reproductivas se manifiesta en los perodos siguientes:
Vaca; de 14-18 anos.
Yegua: alrededor de los 18 anos.
Oveja; entre 6 y 9 aos,
Cerda: de 7-9 aos.
Perra y gata: de 11-13 aos.
Coneja: a los 8 aos.
Gallina: declina la puesta a los 3 aos.
Pubertad
La pubertad se define como el perodo en que el animal alcanza la capacidad de
reproducirse de modo natural. Dicho perodo comienza con la primera ovulacin; en la
novilla esta no se acompaa de manifestaciones de celo y termina cuando la ovulacin se
acompaa de tales manifestaciones.
Si se provee a las novillas una nutricin adecuada, tras el estro de la pubertad se establecen
ciclos regulares [3].
EDAD EN QUE INICIA LA PUBERTAD
En condiciones normales la pubertad empieza alrededor de los 12 meses en el ganado
vacuno; de 15-18 meses en el equino; en la oveja, la cabra y la cerda a los 6 o 7 meses y en
la coneja de 3 - 4 meses.
Las novillas no arriban a la pubertad a una edad o a un peso fijo, pero estos parmetros
interactan para regular el momento del primer estro ovulatorio [4].
Los niveles nutricionales modulan la edad de la pubertad. Si el crecimiento es acelerado
por sobrealimentacin el animal alcanza la pubertad joven. Por otro lado si el crecimiento
es lento por estar subalimentado, la pubertad es demorada [5].
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
La influencia de tres niveles distintos de energa en la dieta, sobre la aparicin del primer
estro, en novillas Holstein, fue objeto de investigacin por Sorensen y otros (Tabla 2.1).
TABLA 2.1 EDAD Y PESO EN EL PRIMER ESTRO EN NOVILLAS HOLSTEIN
ALIMENTADAS CON TRES NIVELES DE ENERGA (SEGN NORMAS DE
MORRISON) [6]
Ingestin
TND
Baja
60 %
Edad
Peso (Ib)
(semanas)
(meses)
Rango Promedio Rango Promedio Rango Promedio
59-80
72
13,6-18,5 16,6
430-575 540
Normal
100%
37-55
49
8,5-12,7
11,3
440-650
580
Alta
140%
29-43
37
6,7-9,9
8,5
460-640
580
En ese estudio qued manifiesto que el peso es fundamental para alcanzar la pubertad, en
las novillas; as como que aumentando el nivel de energa en la racin se puede llegar al
peso necesario a una edad ms temprana.
Las novillas Ceb criadas en sistemas extensivos en las regiones tropicales, tienen una
demora considerable para alcanzar la pubertad, pero se ha comprobado, que si se (es
garantiza una ganancia diaria de 500 g, son capaces de presentar la pubertad a una edad
razonable (Tabla 2.2).
TABLA 2.2 EDAD, PESO Y GANANCIA DIARIA HASTA LA PUBERTAD EN
NOVILLAS CEB [7]
N
Peso (Ib)
Inicio
Edad (meses)
311
Ganancia diaria (g)
7,2
15
Pubertad
500
600
15,4
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
La madurez somtica depende en gran parte del tipo y de la calidad de la alimentacin, por
lo que es mejor orientar el inicio de la reproduccin de las novillas segn su peso. Es decir,
comenzar la inseminacin o cubricin cuando alcanzan tres cuartos del peso promedio de
las vacas adultas de su raza [8].
Cuando los alimentos suministrados a los animales no logran cubrir sus demandas de
energa y protena, se producen trastornos en su comportamiento reproductivo y en la
fertilidad [9].
La alimentacin deficiente conduce al llamado cuadro de seudohipofisectoma, que se
caracteriza por el bloqueo o insuficiencia funcional de la hipfisis. Se observ que novillas
Holstein alimentadas con una racin que contena mayor cantidad de TND y protenas,
tenan hipfisis mayores, por tanto, el incremento del volumen de la hipfisis puede
aumentar la cantidad total de gonadotropina liberada.
En todos los mamferos la malnutricin disminuye la frecuencia de los pulsos de LH, pero
la maduracin del sistema nervioso central (SNC) y, por consiguiente de las gnadas
contina, de modo que, tan pronto se restaura la ingestin de un suministro calrico
abundante, la LH es secretada a un ritmo normal y hay una rpida recuperacin del
desarrollo puberal (Figura 2.1).
Aspectos metablicos en las novillas asociados al sistema de alimentacin
La valoracin del estado metablico de los animales se puede realizar a travs de un perfil
de metabolitos sanguneos, teniendo en cuenta que estos son un reflejo directo del
metabolismo.
La concentracin de los constituyentes sanguneos se regula, en muchos casos, por
mecanismos homeostticos, con la influencia del nivel de absorcin, sntesis o liberacin
de otros de las reservas tisulares.
Cuando la homeostasis falta y las reservas tisulares se tornan inadecuadas, las fuentes
dietticas ejercen una mayor influencia sobre la concentracin de los citados constituyentes
[10].
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
FIGURA 2.1 Efectos de los niveles de nutricin en la oveja sobre la secrecin pulstil de LH. Recuperacin
de los pulsos de LH despus del retorno a una ingestin calrica normal [11]
Perfiles hormonales
Los eventos endocrinos asociados con la pubertad fueron estudiados en novillas de carne
(Bos taurus) por Gonzlez Padilla y otros [12], los que encontraron que los niveles de LH
fluctuaban marcadamente durante el perodo prepuberal, pero presentaban dos picos
caractersticos, con un intervalo entre ellos de 9-11 das, el segundo pico o pico puberal era
seguido de ovulacin e inicio de actividad ovrica cclica normal. Cada pico de LH estuve
precedido de una elevacin en los niveles perifricos de progesterona que duraban de 2-5
das (Figura 2.2).
FIGURA 2.2 Cambios endocrinos asociados con la pubertad [12]
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
Variaciones en otras especies
PUBERTAD EN LA YEGUA
En la yegua la aparicin de la pubertad est en dependencia de la raza y la talla del animal;
adems surte efecto la poca en que se produjo el nacimiento en relacin con la
luminosidad, la alimentacin y el clima.
En general, los primeros celos aparecen entre los 14 y 18 meses con un rango de 10-24. Se
considera que para las razas de talla pequea o razas ligeras, ocurre a los 10-12 meses; para
las de talla mediana o semipesadas a los 15-18 meses y para las de talla pesada a los 18-24
meses.
En todo caso, nunca se debe intentar la cubricin antes de los dos aos para las hembras de
razas ligeras y semipesadas y de los tres aos para las de razas pesadas. Mitchell y Alten
[13] reportaron que de 137 potrancas cubiertas al primer ano de edad concibieron 95 y de
estas abortaron 44 entre los 30 y 60 das iniciales de la gestacin.
PUBERTAD EN LA CERDA
Las lechonatas mantenidas en condiciones idneas de higiene y alimentacin presentan sus
primeros celos a los 6-7 meses de edad, con un peso de 80-90 Kg.; aunque la edad resulta
el factor determinante en la aparicin de la pubertad en la cerda, se ha comprobado que las
hembras mestizas (cruces industriales) se adelantan en 15 o ms das a las razas puras.
Asimismo las nacidas en invierno son ms precoces que las nacidas en verano.
La cubricin se debe realizar alrededor de los nueve meses cuando los rganos genitales
sufren un desarrollo acelerado (cuatro veces mayor que al inicio de la pubertad) y con un
peso corporal que sobrepasa los 100 Kg.
PUBERTAD EN LA OVEJA Y LA CABRA
Independientemente que la edad desempee un papel preponderante en la presentacin de
las primeras inquietudes sexuales en estas especies, estn sujetas tambin a la poca del
nacimiento, en relacin con la luminosidad, al igual que todos los animales fotoperidicos,
en este caso de das cortos.
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
Las hembras que arriben a los 6-8 meses de nacidas, en las zonas templadas, al comenzar
la estacin o poca de cubriciones (menor incidencia de la luz solar), ya presentan celos
puberales, aunque con muy pocas manifestaciones externas. Las nacidas tardamente con
respecto a la poca de cubriciones, en dichas zonas templadas, no manifiestan celo hasta
aproximadamente los 14 meses de edad.
Al igual que en las novillas, la aparicin de la pubertad en las hembras, tanto de ovejas
como de cabras est influida por su peso corporal, no presentndose hasta alcanzar ms del
60 % del que tendrn en la etapa adulta. En todo caso estas hembras no deben ser cubiertas
hasta despus del primer ano de vida, aun en nuestras condiciones subtropicales donde la
manifestacin de los celos puberales ocurre alrededor de los 8-10 meses y, prcticamente,
en cualquier poca del ao.
CICLO ESTRAL
El ciclo estral est representado por un complejo de transformaciones hormonales,
histolgicas y morfolgicas, que actuando bajo influjos neurohormonales no solo crean el
cuadro a nivel de los rganos reproductores sino que tambin afectan otros rganos de la
economa.
La finalidad de la actividad cclica estral es preparar las condiciones favorables para la
fecundacin, nidacin y desarrollo del feto.
Los ciclos son el resultado de la correlacin de factores hereditarios y ecolgicos, donde
los primeros dependen de los segundos, en forma tan intensa que no solo dominan el ciclo,
sino toda la esfera sexual [14].
En algunas especies los ciclos se suceden uno tras otros, son las especies de ciclo continuo;
en otras la actividad estral solo se manifiesta en determinados perodos, estas son las de
ciclo estacionario. Como ejemplo de las de ciclo continuo se halla la vaca y la cerda y de
estacionario la yegua, la oveja y la cabra.
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
Las de ciclo estacionario pueden presentar estos en la poca del ao en que los das son de
corta duracin, especies fotoperidicas de das cortos, como la oveja y la cabra; o por el
contrario presentarlos en la poca en que los das son de larga duracin, fotoperidicas de
das largos como la yegua. Esta caracterstica fotoperidica es bien manifiesta cuando nos
separamos del Ecuador, ganando en altitud y tiende a disminuir cuando nos aproximamos.
Si los animales fotoperidicos de un hemisferio terrestre son trasladados al otro, mantienen
la caracterstica fotoperidica de sus ciclos, invirtiendo por tanto los meses del ao en que
los presentan.
Clasificacin de las especies segn la repeticin del ciclo estral.
Poliestricas o policiclicas continas
Rumiantes y cerda
Poliestricas estacinales de das largos
Yegua y gallinas
Poliestricas estacinales de das largos
oveja
Diestricas
Perra y otros carnvoros
Potenciales o inducidas
Roedores
Duracin del ciclo estral
La duracin del ciclo estral, as como la de sus fases, vara en las distintas especies (Tabla
2.3).
TABLA 2.3 DURACIN DEL CICLO ESTRAL [15]
Especies
Vaca
Duracin (das)
21-22
Yegua
19-25 (con gran variacin de acuerdo con la duracin del celo)
Cerda
19-20
Oveja
16-17
Cabra
21 (tambin ciclos cortos)
En la vaca el ciclo estral tiene una duracin media de 21 das. En la raza Ceb se observ,
en un anlisis de 229 ciclos presentados por 15 novillas a partir de su pubertad, que sus
primeros siete ciclos tuvieron una duracin media de 17,4 das, mientras que los
posteriores (127 ciclos) duraron 21,7 das [7].
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
Fases del ciclo estral
El ciclo estral se divide en cuatro fases que son: estro, metestro, diestro y pro-estro (Figura
2.3).
FIGURA 2.3. Ciclo estral en los animales y sus alternativas [16]
Estro o celo. Esta es la fase ms significativa del ciclo estral, marcando su inicio (da 1 del
ciclo) y dndole nombre. Se caracteriza por manifestarse la libido, con disposicin de la
hembra a aceptar la monta. En esta fase se produce la ovulacin, excepto en la vaca, en la
que ocurre entre 10-12 h despus de terminado el estro [17].
Metestro. En esta fase se inicia la actividad progesternica, con la formacin del cuerpo
lteo. Durante ella se desarrolla la gestacin si la hembra es fecundada. De no ocurrir la
fecundacin a esta fase le sucede un perodo de quietud sexual, el diestro.
Diestro. Constituye una fase de reposo de las funciones estrales con franco predominio de
la funcin luteal.
Pro-estro. Se caracteriza por el inicio de la actividad folicular, al desaparecer el dominio
del cuerpo lteo.
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
De acuerdo con la actividad hormonal ovrica que prevalezca, el ciclo se puede dividir en
una fase folicular o estrognica (pro-estro -estro) y una fase progesternica (metestrodiestro) [18].
Manifestaciones clnicas del celo en la vaca
La irritacin nerviosa provocada por los niveles estrognicos elevados, en la vaca, hace
que se modifique su. comportamiento habitual: intranquilidad, molestan a las otras vacas,
mugen con frecuencia, toman actitud de bsqueda del macho, estn inapetentes,
disminuyen su produccin lctea, que sufre cambios fsico-qumicos lo que le brind la
leche un sabor salino y motiva su coagulacin al hervirla (variacin en la. composicin
proteica). Se nota tambin: un reblandecimiento de las heces relacionado con la actividad
de movilizacin inica de la foliculina.
El cuadro sintomtico externo se complementa con las modificaciones que se aprecian en
la vulva, esencialmente su aspecto edematoso, con prdida de su sistema plegado,
prolongacin de la rima que entreabre los labios vulvares y, adems, muestra una marcada
hiperemia en la mucosa. Pende de este rgano el tpico moco estral, transparente, filante,
que gana en densidad hacia el acm del celo, donde se torna ms consistente. Estas
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
secreciones embadurnan la vulva, el maslo de la cola, los corvejones, y toda el rea aledaa
al genital externo.
Al relajarse los ligamentos anchos de la pelvis (sacro espinoso y sacro tuberoso) dan la
apariencia del sacro elevado y la cola hundida.
Se observan los movimientos rtmicos del ano, provocados por las contracciones
musculares del vestbulo vaginal, el elevador del ano y el esfnter anal externo.
En todo caso siempre que se detecte una hembra intentando montar a otras hembras,
realizando la mmica sexual, y permitiendo que otras hembras monten sobre ella (actitud
de bisexualidad), se puede afirmar que est en celo, ya que constituye la manifestacin
tpica de la vaca en la fase del ciclo estral.
Se debe tener en cuenta que, en ocasiones, los sntomas externos del celo suelen verse
atenuados o faltar completamente, celos silentes, los que requieren del toro recelador o de
la exploracin de los rganos genitales, por el mtodo rectovaginal para su deteccin.
Por lo regular, las hembras que participan montando a las que son recogidas en celo, se
encuentran en pro-estro, siendo detectadas a su vez a los 2 o 3 das, lo cual establece un
ritmo que se debe considerar para una mayor eficiencia en la actividad del personal
dedicado a esta labor.
Acorde con los cambios ya descritos en el comportamiento de la hembra, el estro evidencia
una serie de manifestaciones en los rganos genitales internos, que se puedan apreciar a
travs del recto (ovarios, cuernos) o mediante el especulum (vagina, vestbulo vaginal,
cervix).
Especial importancia tiene la valoracin de los cambios que se suceden en los genitales
internos, cuando se aplica la tcnica de la inseminacin artificial como mtodo zootcnico
de reproduccin. Aunque la palpacin ovrica no es aconsejable en la hembra que va a ser
servida, se puede notar en este rgano la presencia de un folculo desarrollado (2 cm de
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
dimetro aproximadamente) como una vescula cuya fluctuacin se acenta prximo a su
dehiscencia.
Valiosas resultan las modificaciones que se evidencian en los cuernos uterinos, los que se
encuentran contrados, aprecindose sus curvaturas mayor y menor (semejan las astas de
un carnero montas) dadas por el incremento de la actividad estrognica sobre la
musculatura lisa uterina (tono) con consistencia aumentada. Estas caractersticas de tono y
consistencia son ligeramente apreciables en el pro estro, alcanzan su mxima expresin en
la misma medida en que el celo se desarrolla, declinando en el metestro.
Las manifestaciones externas del celo pueden faltar (celo silente), pero siempre estarn
presentes los cambios internos. En la prctica de la inseminacin artificial es
imprescindible dominar estos aspectos para valorar el momento ptimo en que esta debe
realizarse, en relacin con la ovulacin.
A la exploracin vaginal se observa la tpica hiperemia, edematizacin y relajacin de todo
el canal vaginal que le brinda a la mucosa un color rosado intenso y un brillo peculiar. El
fondo vaginal dilatado acumula secreciones procedentes de la vagina y el crvix, el que se
muestra edematoso, hipermico, y pierde su caracterstica forma cnica plegada, dilatado
en forma aplanada y con los pliegues engrosados; el canal cervical se muestra entreabierto.
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
Sintomatologa clnica del metestro.
El metestro es una etapa de transicin de la fase estrognica a la lutenica. Se caracteriza,
en general, por la declinacin, hasta desaparecer, de los sntomas externos del celo y
cambios en los rganos genitales internos que culminan la preparacin para la recepcin y
el desarrollo de un nuevo individuo, y esta se anula en caso de no haber fecundacin. En la
vaca ocurre la ovulacin en los inicios de esta fase.
Se puede notar el cese de la congestin vulvar, al ceder el cuadro edematoso y disminuir la
cantidad del moco estral que se hace denso y opaco; en la vaca (50 %) y las novillas (90%)
aparecen algunas estras sanguinolentas en estas secreciones postestrales, como resultado
de la diapdesis en los capilares sanguneos, rotos al final del estro, cuando se cambia la
fase folicular (proliferativa) en progesternica (secretora).
Lo ms significativo que ocurre a nivel ovrico en esta fase es la formacin del cuerpo
lteo. El metestro dura en la vaca de 2-3 das. Los cuernos uterinos pierden su tono y
aparece una flacidez progresiva; su consistencia disminuye porque cesa la actividad
estrognica. La vagina se torna plegada, la hiperemia es menos intensa con pocas y densas
secreciones de color opaco; la flor radiada se constrie cerrando el conducto cervical.
Sintomatologa clnica del diestro
Esta etapa de reposo sexual se caracteriza por prevalecer la actividad progesternica por
parte del cuerpo lteo. En la vaca, a los 5-6 das del ciclo se palpa con seguridad el cuerpo
lteo. En la yegua es palpable a los 4 - 5 das en la fosa ovrica y su evolucin contina
dentro de las estructuras ovricas, no aflorando al exterior. Su deteccin es imposible
despus de los ocho das posteriores a la ovulacin.
En esta fase y hacia el da 16 del ciclo, el cuerpo lteo sufre un proceso lteo ltico.
Sintomatologa clnica del pro-estro
El pro-estro es la ltima fase del ciclo y es una etapa de preparacin para el estro. Se
caracteriza por un incremento de la estrognesis, al terminar el predominio de la
progesterona que produca el cuerpo lteo. El incremento de estrgenos se traduce en
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
ligeras modificaciones de la conducta de la hembra; inapetencia, intranquilidad, hasta
intentar montar a otras hembras.
Se inicia una congestin y tumefaccin ligera de la mucosa vaginal, en la que comienza
una secrecin mucosa acuosa, hasta que se hacen tpicas las manifestaciones de celo y, con
ello, el comienzo del nuevo ciclo.
Manifestaciones clnicas del celo en otras especies
CELO EN LA YEGUA
Se reconoce el celo en la yegua por la tumefaccin vulvar y las descargas de moco
abundante, siendo tpica la presentacin del reflejo de Blitsen o centelleo que consiste en
la exposicin peridica del cltoris y la emisin de cortos chorros de orina, Se comporta
intranquila, nerviosa, da frecuentes relinchos y coceos, escarba con los cascos, mirada atrs
hacia los cuartos traseros, al tiempo que fustiga sus flancos con la cola y mantiene un
activo amusgamiento.
La comisura bucal de la yegua en celo est elevada y hacia atrs; el labio inferior muestra
un aspecto tpico, como de quien atrapa algo, en tanto que los ojos muestran una mirada
peculiar fija. A estos sntomas se les llama cara de celo.
El ritual del celo comprende una seria agresividad hacia el macho, lo que obliga a su
proteccin, empleando para ello atalajes adecuados (trabones) que limitan los movimientos
de la hembra. Puede reconocerse la inminencia de la ovulacin por la mayor disposicin
receptiva de la yegua, adems de que las descargas mucosas pasan de viscosas y claras a un
estado ms acuoso; pasada la ovulacin se hacen ms viscosas, consistentes y de
coloracin griscea.
La valoracin de los cambios en los rganos genitales ayudan a pronosticar el momento de
la ovulacin, Tiene especial inters el cuadro vaginal y el ovrico, que muestra una
prolongacin de la teca interna folicular en forma de cua (teca interna cnica) y puede ser
palpada prxima a la fosa de ovulacin, adems, se aprecia una amplia vascularizacin peri
ovrica propia del estro. Cerca de la ovulacin el folculo se torna flcido y blando, de 4-5
cm de dimetro.
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
En la vagina se puede observar un color rosado intenso, mucosa brillante y gran cantidad
de moco claro en la porcin anterior. Debe tenerse en cuenta que la mucosa comienza a
congestionarse al ser expuesta al aire, por lo que despus de la introduccin del espculo
muestra inmediatamente, su cambio de color.
La abertura externa del crvix, al inicio del celo, es cilndrica, hinchada, edematosa y los
pliegues cervicales se relajan; estos sntomas avanzan hasta que prximo a la ovulacin el
hocico de tenca suave al tacto se torna rgido y con contracciones rtmicas; lo mismo
sucede al contacto peneano.
Como ya ha sido descrito, el ritmo estral en la yegua obedece a los cambios de la
iluminacin solar. Al inicio de la poca de cubriciones el celo se extiende hasta 2-3
semanas y se regula a 5-7 das paulatinamente; el nuevo celo se inicia alrededor de los 16
das de haber finalizado el anterior, pudindose afirmar que este determina la duracin del
ciclo estral.
CELO EN LA OVEJA Y LA CABRA
En general el cuadro sintomtico del celo recuerda al de la vaca, pero mucho ms
atenuadas las manifestaciones externas (muy frecuente el celo silente en estas especies).
No hay un marcado inters en la bsqueda del macho, lo que obliga al uso de receladores
vasectomizados o con el pene desviado, a estos se les aplica cualquier sustancia colorante
(preferiblemente en la regin esternal), de forma tal que deje las huellas de que mont a la
hembra.
La ligera inquietud, los balidos, la cola ligeramente alzada, con cortos y rpidos
movimientos y la vulva tumefacta, son indicios del celo en la hembra, que dura de 30-40 n.
CELO EN LA CERDA
A diferencia de las otras especies ya descritas, la cerda asume una actitud pasiva en el
momento del celo. Grue y hociquea otras hembras de la piara, se comporta inapetente,
busca al macho y la vulva sufre de una amplia tumefaccin e hiperemia. Ya en presencia
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
del macho asume el tpico reflejo de la inmovilidad, que puede ser aprovechado por el
hombre en la deteccin del celo; manifiesta esta quietud cuando nos apoyamos en sus
ancas, preferiblemente a horcajadas.
CELO EN LA PERRA
Bajo la influencia de niveles incrementados de estradiol, la perra durante el pro-estro
comienza a ser atractiva para los machos. Hay un aumento en el tamao y la turgidez de la
vulva y el rea perineal, y una descarga vaginal serosanguinolenta debida a la presencia de
hemates que escapan del congestionado endometrio. Los frotis vaginales estn
compuestos principalmente de clulas epiteliales redondas y poligonales [19].
El pro-estro tiene una duracin de una semana, pero puede reducirse a solo tres das o, por
el contrario, prolongarse hasta tres semanas.
La aceptacin a ser montada por el macho, o estro, ocurre por lo general despus de la
descarga de LH que dura 2-3 das. Los niveles de progesterona se incrementan en el da del
pico de LH y los das posteriores (Figura 2.4). El flujo sanguneo cesa o disminuye
mientras que la tumefaccin vulvar alcanza su mximo.
FIGURA 2.4 Promedio de niveles de estrgenos. LH y progesterona durante el pro-estro y estro de perras
Beagle y promedio de Duracin de diferentes signos clnicos [19]
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
La ovulacin ocurre en un momento impredecible que puede variar desde 2 o 3 das
anteriores a 4 o 5 das posteriores al Comienzo del celo.
CELO DE LA GATA
El celo se produce por un aumento de 17p estradiol en el plasma de 20-25 pg/mL en el
proestro a 35-70 pg/mL. Menos del 10 % de las gatas muestran signos de estro el primer
da del incremento hormonal, para el tercer da, 50 % de ellas presentarn signos de estro.
En la gata no hay cambios de la apariencia de los genitales externos durante el celo, solo
hay cambios de comportamiento. Disminuye su actividad general, maullan, avanzan con
sus patas traseras y se frotan contra objetos. Ellas aceptan ser montadas y entonces en el
estro ser cubiertas. El celo dura alrededor de 10 das y en las gatas Persas menos de 6 das.
Los celos largos de ms de 19 das al parecer corresponden a una repeticin de celo, en la
que el diestro no fue observado [20].
Las gatas rehsan ser montadas las 2-3 semanas posteriores al celo (diestro).
MODIFICACIONES ESTRUCTURALES DEL TRACTO GENITAL
En el transcurso del ciclo estral, cada rgano del aparato genital (ovarios, oviducto, tero,
cuello, vagina) sufre modificaciones cclicas, con un ritmo de correspondencias e
interacciones entre ellos, lo que posibilita el logro de los objetivos biolgicos de esta
actividad orgnica: la perpetuacin de la especie.
Ciclo ovrico
El ovario presenta una estructura medular muy vascularizada e inervada, y otra perifrica o
cortical de color grisceo, compuesta bsicamente por los folculos ovricos y el tejido
intersticial; la masa ovrica est cubierta por la tnica albugnea (tejido colgeno), y esta, a
su vez, por una capa de clulas cbicas (epitelio germinativo).
En los quidos aparece la tnica albugnea sumamente gruesa y con la bolsa ovrica
adherida de manera directa sobre el rgano, lo que le da una consistencia ms firme y una
mayor resistencia que en el resto de las especies; a nivel ventral hay una depresin (fosa de
ovulacin) menos resistente y libre de tejido fibroso, que permite la proyeccin del vulo,
la que en esta especie solo es posible por este sitio.
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
Esencialmente el curso de la actividad ovrica se encamina al desarrollo del vulo y de las
estructuras foliculares, para culminar con la formacin y la evolucin del cuerpo lteo, los
cuales determinan una fase folicular o estrognica y otra luteal o progesternica.
OVOGNESIS
Las ovogonias se producen durante la vida fetal por multiplicacin mittica de clulas
germinales, que se multiplican hasta que la ltima generacin pasa a la profase de la
primera divisin meitica, punto en el que son ovocitos primarios. El ncleo del ovocito
primario se mantiene en la etapa de diacinesis o reposo de la citada profase hasta que el
folculo alcanza la madurez en la etapa preovulatoria.
Se supone que un factor peptdico, llamado inhibidor de maduracin del ovocito (OMI) es
el encargado de mantener al ovocito en la mencionada fase de diacinesis [21]. En la vaca
este permanece en la fase de paquinema y en la cerda en la de diplonema [22].
La ternera nace con un cierto nmero de ovocitos primarios, el que vara grandemente,
calculndose de unos 40 000-800 000 [23]. Estos disminuyen muy rpido a partir del
nacimiento y despus de este lo hacen de forma gradual.
FOLCULOS PRIMORDIALES
Los folculos primordiales estn constituidos por ovocitos primarios rodeados de una capa
simple de clulas ms pequeas, aplanadas (epitelio folicular), que darn origen con
posterioridad a las clulas de la granulosa. Se encuentran situados en la zona cortical del
ovario, debajo de la tnica albugnea.
DESARROLLO FOLICULAR
La folculognesis es un proceso dinmico y rpido, que se produce durante el pro-estro y
el estro, y consiste en la formacin de un folculo de De Graaf o folculo maduro, a partir
da uno primario (Figura 2.5).
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
FIGURA 2.5. Esquema del desarrollo folicular.
En el transcurso de cada ciclo de actividad ovrica, una serie de folculos comienzan su
desarrollo, la mayora de los cuales no llegan a madurar y sufren atresia. El folculo
destinado a ovular recibe mayor cantidad de sangre, y, adems, sus capilares son ms
permeables que los de los restantes folculos [24].
El folculo ovrico proporciona al ovocito un medio apropiado para su maduracin, el que
en algunas especies (mujer y coneja) se ha determinado que incluye una temperatura de 2-3
C ms baja que su temperatura sistmica [2]. Al comenzar su crecimiento se produce la
migracin hacia la parte ms profunda del ovario.
El desarrollo folicular se inicia con el aumento del nmero de clulas que rodean al
ovocito, las cuales forman una nueva capa a su alrededor, folculo secundario. Durante el
crecimiento de este, se deposita una capa glicoprotenica compleja, entre la membrana
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
celular del ovocito y las clulas foliculares que lo rodeaban, que va a formar la zona
pelcida, membrana clara, delgada no celular, que se engruesa al incorporar sustancias
producidas por las clulas foliculares y, posiblemente tambin, por el propio ovocito
(Figura 2.6). Cuando el crecimiento de este ha terminado, las clulas foliculares comienzan
a separarse y aparecen cavidades que se van llenando de lquido (liquor folliculi) fluido
viscoso y rico en sustancias secretadas por las clulas de la granulosa.
Las citadas cavidades confluyen para formar una cavidad nica, el antrum folicular. Las
clulas foliculares que forman la pared de este antro, se denominan clulas de la granulosa.
El folculo, llamado entonces folculo cavitario, crece como consecuencia del aumento de
lquido folicular que empuja y comprime los tejidos circundantes. (Figura 2.7).
A medida que contina este crecimiento, el ovocito queda localizado cada vez ms
excntricamente, rodeado por un acumulo de clulas de la granulosa, el cumulus oophorus.
(Figura 2.8) La capa de clulas que rodea inmediatamente al ovocito, formndole una
especie de anillo, es denominada corona radiada.
FIGURA 2.6 Ovocito con su corona radiada
Formada por clulas de la granulosa .
Durante el crecimiento folicular, las clulas del tejido conectivo que lo circundan, se
diferencian como una tnica, la teca folicular, la que est compuesta por una tnica interna,
celular y vascular y por una externa fibrosa [25]. Las clulas de la teca interna estn
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
separadas de las de la granulosa folicular por la membrana propia. Coincidiendo con la
aparicin de la teca folicular se produce el ascenso del folculo a la superficie del ovario.
Figura 2.7
Figura 2.8
Ondas de desarrollo folicular
Hasta los estudios de Rajakoskl [26] y [27] se supona que la curva de crecimiento
folicular comenzaba a mediados del ciclo estral y progresaba constantemente hasta que se
iniciaba la maduracin preovular, pero este, investigando las variaciones del nmero de
folculos en crecimiento, de novillas sexualmente maduras, sacrificadas en diferentes fases
del ciclo estral, observ que dicha curva se produca en ondas, interpretando que estas
ondas eran dos, una comenzaba entre el tercer y el cuarto da del ciclo y la otra entre el
decimosegundo y el decimocuarto da, que ambas daban origen a un folculo de tamao
preovular y que el folculo grande de la primera onda de crecimiento sufra atresia desde el
decimosegundo da.
Posteriormente se encontr que el crecimiento folicular se puede producir en dos o tres
ondas [17] y [28], cada una con un folculo dominante, as como que el ltimo de estos es
el nico que llega a ovular. La primera onda se inicia despus de la ovulacin y su folculo
dominante alcanza su mayor tamao al final de la primera semana del ciclo y sufre atresia;
la segunda onda se produce a mediados del ciclo y su folculo dominante alcanza su mayor
tamao cerca del fin de la segunda semana del ciclo, e igualmente sufre atresia y la tercera
y ltima onda de crecimiento folicular se origina cerca del final del ciclo y su folculo
dominante madura, y ovula poco despus de terminado el celo.
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
El avance de la tecnologa de observacin con ultrasonido, con su impresionante capacidad
de seguir la secuencia de crecimiento y atresia o de ovulacin de los folculos dominantes,
ha mostrado claramente que las novillas pueden tener 2-3 folculos dominantes durante el
ciclo [29].
Cuando el ciclo dura de 18-21 das ovula el segundo folculo dominante y cuando el ciclo
se prolonga hay una tercera onda de maduracin folicular y ovula el tercer folculo
dominante. Esto sugiere que el tiempo que demore en regresar el cuerpo lteo desempea
un papel importante en determinar si el ciclo tendr dos o tres ondas de maduracin
folicular [29].
Sobre el da 18, invariablemente estn presentes dos grandes folculos, pero el dominante
tiene una cantidad de estradiol mucho mayor que el otro, destinado este a sufrir atresia
[30].
Los folculos dominantes por tener una mayor afinidad por las gonadotropinas fijan estas
hormonas en mayor cantidad que los folculos menores y secretan la mayor parte de los
estrgenos producidos por los ovarios, con lo que impiden contine el desarrollo del resto
de los folculos.
Solo el folculo dominante de la ltima onda del ciclo ovula, porque los altos niveles de
progesterona que se mantienen durante el resto del ciclo bloquean el pico preovulatorio de
la hormona luteinizante (LH) [31].
Las vacas que tienen en su posparto ciclos cortos de 8-13 das de duracin, en estos ovulan
su primer folculo domnate [21].
Atresia folicular
El destino final de la mayora de los folculos es la atresia. Estos la pueden sufrir en
cualquier estadio de su desarrollo. Cuando un folculo entra en una onda de maduracin
ovula o sufre atresia. Adems, los folculos que no estn en la onda de crecimiento tambin
pueden degenerar [30].
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
En los folculos de las perras se pueden distinguir dos patrones de atresia. Un tipo
predomina en los folculos secundarios, en los que prevalecen los cambios necrticos en
los ovocitos y zonas pelcidas, y donde las alteraciones de las clulas de la granulosa son
secundarias, y otro tipo que es el nico encontrado en los folculos terciarios, que se
caracterizan por cambios en la granulosa y sin casi ninguno en el ovocito y la zona
pelcida [12].
En los grandes folculos, la atresia se puede reconocer por la presencia de cuerpos
pignticos (material nuclear condensado), cuyo nmero determina su estadio. A medida
que el nmero de estos cuerpos se incrementa, la capa granulosa desaparece y las tecas
cambian a tejido intersticial.
El ltimo estadio de la atresia es la invasin de la cavidad folicular por fibroblastos y la
desaparicin del tejido intersticial. El ovocito degenerado o solo la zona pelcida son las
ltimas estructuras que se pueden ver en un folculo al final de la atresia [32]. En casos
patolgicos los folculos son hemorrgicos o qusticos.
DIVISIN MEITICA
Al terminar el crecimiento folicular, casi 3 h despus de producirse el pico preovulatorio de
LH, el ncleo del ovocito que haba permanecido hasta ese momento en la etapa de reposo
de la profase de meiosis, reinicia las divisiones meiticas. Durante la primera divisin el
ncleo se acerca a la superficie celular y pierde su membrana, desaparecen los nuclolos y
acontece un hecho de importancia capital, la mitad de sus cromosomas y una pequea
cantidad de citoplasma es expulsado, los que forman una clula minscula, el primer
corpsculo polar o polocito, el que va a situarse en el espacio perivitelino.
Esta divisin de reduccin cromsomtica, proceso denominado de maduracin nuclear, se
completa en casi todas las especies domsticas, pocas horas antes de la ovulacin [9] y da
lugar a un ovocito secundario, que tiene un nmero haploide de cromosomas. En la perra y
en algunas yeguas, esta primera divisin meitica tiene lugar despus de la ovulacin.
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
La segunda divisin meitica se produce en el salpinx, cuando un espermatozoide penetra
en la zona pelcida del ovocito secundario y lo activa. Esta divisin es ecuacional, y no
difiere en su esencia de una mitosis comn [25], dando lugar a la formacin de un segundo
corpsculo polar, que va a situarse igualmente en el espacio perivitelino y a un vulo
maduro u ovtide. En algunos animales, el primer corpsculo polar tambin se divide, por
lo que el ovtide puede contener tres corpsculos polares bajo la zona pelcida.
El vulo de todas las especies domsticas, en el momento de la ovulacin, es una de las
clulas ms grandes del organismo, variando de 0,12-0,185 mm de dimetro, sin incluir la
zona pelcida; este crecimiento que triplica su tamao, si es fertilizado, asegura la
nutricin durante sus primeras divisiones.
OVULACIN
En todos los mamferos la ovulacin u ovocitacin se produce al romperse el folculo de
De Graaf por una zona vascular de su superficie denominada estigma, liberando al ovocito
hacia el oviducto, donde completa su maduracin. La ovulacin ocurre en cualquier punto
de la superficie del ovario, excepto por el hilio.
La yegua tiene la particularidad, de que en ella la ovulacin nicamente se origina en una
zona delimitada de su ovario llamada fosa de ovulacin. En dependencia del tamao del
estigma y de la localizacin del cumulus oophorus, la ovulacin puede requerir solo
algunos segundos o durar muchos minutos [221.
La ovulacin nada ms ocurre en animales pberes no gestantes, durante los ciclos estrales.
No obstante, las yeguas frecuentemente ovulan al principio de la gestacin [33] y la vaca
puede ovular tambin mientras transcurre la gestacin, pero esto sucede muy raramente. En
muchas especies la ovulacin es espontnea, pero en la gata, la coneja y la hurona, es
provocada por el coito.
Aproximadamente se producen en el ovario derecho el 60 % de las ovulaciones en la vaca
y en la oveja el 52-59 %. Sin embargo, en la yegua y en la cerda esta actividad ovrica es
de un 61 % y un 55-60 % respectivamente. En la vaca, la yegua y la cerda la ovulacin se
manifiesta al azar con respecto al ovario que contiene el cuerpo lteo del ciclo anterior.
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
Para que se produzca la ovulacin es necesario que se rompan todas las capas celulares que
separan al ovocito del exterior del folculo, estas son: el epitelio superficial del ovario, la
tnica albugnea, la membrana basal y el estrato granuloso.(Figura 2.9)
Como resultado de la despolimerizacin de los muco polisacridos que conforman la pared
folicular, se observa un debilitamiento de las capas externas de esta estructura, lo que les
permite proyectarse en forma de cono y origina la papila ovulatoria, estigma, la que se
adelgaza y agrieta permitiendo la salida del licor folicular, los restos foliculares, el ovocito
y sus cubiertas.
Las clulas del cmulo prolgero segregan glucoprotenas, las que forman una masa
viscosa que rodea al ovocito y su corona. Despus de la ruptura folicular esta masa viscosa
se disemina en la superficie del ovario y facilita la captacin del ovocito por las fimbrias,
cuyas clulas ciliadas se encargan de impulsarlo hacia la luz del oviducto [23]. Al mismo
tiempo las clulas de la granulosa se disocian por completo en el pice folicular y al final
desaparece su capa.
Figura 2.9
Mecanismo de la ovulacin
Se han elaborado distintas hiptesis para tratar de explicar el mecanismo de la ovulacin.
Hace aos se pens en la intervencin de un aumento de la presin intrafolicular, pero en la
medida en que se han realizado determinaciones de la presin intrafolicular, se ha ido
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
comprobando que no hay aumentos de presin antes de la ovulacin; es ms, en la vaca, en
la yegua y en otras especies, la fina pared folicular que durante el pro-estro y principios del
estro usualmente es tensa y firme, es ms flcida y suave precisamente antes de que se
produzca la ovulacin [22]. Hoy da se sabe que aunque el volumen folicular aumenta
rpidamente en las horas prximas a la ovulacin, el folculo no experimenta ningn
incremento en su presin intracavitaria debido a que aumenta su elasticidad. Esto es
resultado de establecerse una cohesin ms laxa de las clulas de la teca externa, causada
por el edema invasor de esta capa y la disociacin de fibras colgenas [34].
Al inicio del estro, cuando los niveles de progesterona en sangre caen a su mnima
expresin y los estrgenos alcanzan sus valores mximos del ciclo estral, una sbita
secrecin de gonadotropina provoca el llamado pico preovulatorio de LH. Esta hormona
induce la produccin de prostaglandina y de esteroides, esenciales al proceso de ovulacin,
pues su inhibicin impide la ovulacin [35].
La prostaglandina F contribuye a la ruptura de los lisosomas de las clulas epiteliales en el
pice folicular. Sus aumentos tambin incrementan las contracciones de las clulas de
msculo liso del estroma y de la teca externa de los folculos preovulatorios, ricamente
inervados por terminaciones del sistema nervioso autnomo [34]. Las contracciones
ovricas facilitan la ruptura folicular e intervienen en la expulsin del ovocito. Los
bloqueadores B adrenrgicos, como el propanolol y el atenolol retrasan la ovulacin. Se ha
demostrado que la FSH tambin induce ovulacin [32].
Momento de la ovulacin
En todas las especies en que la ovulacin es espontnea, usualmente ocurre durante el
estro, con excepcin de la vaca, en la que se produce aproximadamente 11 h despus de
terminado el perodo de receptividad o verdadero celo, o sea, alrededor de 30 h despus de
su comienzo. En las novillas es 2 o 3 h antes que en las vacas. La cpula adelanta la
ovulacin, adems de disminuir la duracin del celo. Novillas cubiertas por un toro
vasectomizado ovularon 7,7 h despus de terminado el celo, mientras que las que no fueron
montadas lo hicieron a las 9,9 h [22] y [36].
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
La yegua ovula de 24-48 h antes de terminar el celo, la oveja de 12-24 h y la cabra y la
cerda prximo a su final. La perra ovula al principio del estro, pero ovocitos que an no
han expulsado el primer corpsculo polar (ovocitos primarios), por lo que no pueden ser
fertilizados hasta alrededor del cuarto da del estro [22] (Tabla 2.4 y Figura 2.10).
FIGURA 2.10 Momento de la ovulacin en distintas especies, sealada por una flecha.
TABLA 2.4 MOMENTO DE LA OVULACIN
Especies
Vaca
Momento estimado
Aproximadamente 11 h despus del celo
Yegua
24-18 h antes de terminar el celo
Cerda
Prximo al final del celo
Oveja
Prximo al final del celo
Cabra
Prximo al final del celo
Perra
Principio del celo
En los animales de ovulacin provocada, esta se produce, aproximadamente 10 h despus
de la cpula en la coneja y en la gata a las 27 h despus de realizada esta. En estas especies
la ovulacin se puede provocar mediante estmulos en la vagina que simulen al coito. La
aplicacin endovenosa de 25 UI de LH a conejas, les provoc la ovulacin entre 101/2 y
10 % h al 50 % de ellas [22]. Este tratamiento puede ser aplicado a las gatas para
terminarles el celo.
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
Desarrollo del cuerpo lteo
Como consecuencia de la ovulacin se produce una hemorragia en la cavidad folicular, la
sangre all depositada se coagula y recibe entonces la denominacin de cuerpo
hemorrgico. Este es infiltrado por eosinfilos y las clulas de la granulosa y las tecales
proliferan rpidamente al utilizar los nutrientes que le proporciona el cogulo y se
luteinizan para formar el cuerpo lteo. Las clulas de la granulosa en 3-5 das alcanzan 20
de dimetro [34].
Cantidades considerables de progesterona se producen y se liberan, a partir del da 3 o 4
del ciclo, del cuerpo lteo, el que alcanza su peso mximo y, con ello, su produccin
mxima de progesterona alrededor del da 11 de dicho ciclo cuando mide
aproximadamente 2 cm de dimetro y pesa cerca de 5 g, superando el tamao del folculo
que le dio origen y en algunas ocasiones es mayor que el resto del ovario. Este es uno de
los crecimientos ms rpidos que se conoce en biologa [25].
Usualmente, el cuerpo lteo hace una prominencia en la superficie del ovario, conocida
como corona del cuerpo lteo y en su superficie se puede distinguir una depresin, que
seala el punto donde se produjo la ovulacin (Figura 2.11).
El cuerpo lteo de la vaca debido a un pigmento amarillo lipocrmico, la lutena, tiene
hacia el sptimo da del ciclo un color caf claro a oro, que cambia a amarillo dorado hacia
el da 14 y de este da al 20 cambia progresivamente al rojo [2].
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
FIGURA 2.11 Obsrvese en el ovario un folculo y un cuerpo lteo.
Si el animal no ha sido fecundado alrededor del da 16 del ciclo, comienza la regresin del
cuerpo lteo, el que disminuye rpidamente de tamao, al ser sustituidas sus clulas
lutenicas por clulas fibrosas, tomando un color ladrillo claro (rubrum). Este cuerpo lteo
no funcional, avascular, en degeneracin, dura dos o tres semanas. Una estructura vestigial
plida, el cuerpo albicans o cuerpo blanco, est a menudo presente durante los dos o tres
ciclos posteriores [37].
Cuando el vulo es fecundado, el cuerpo lteo persiste como cuerpo lteo de la gestacin,
y su funcin hormonal protege el curso de la gestacin. En los casos en que no se produce
la fecundacin, es llamado cuerpo lteo peridico, de ciclo o falso [8].
Ciclo tubrico
Los oviductos constituyen la va de comunicacin del tero con los ovarios, de ah que su
actividad funcional tenga gran importancia, ya que el pabelln es el rgano de captacin
del vulo cuando se produce la ovulacin y la ampolla tubrica es el sitio donde se
encuentra el vulo con los espermatozoides y tiene lugar la fecundacin (anfimixis).
Adems, es el sitio donde transcurren las primeras horas del cigoto.
Los oviductos poseen una mucosa, una muscular y una serosa. La mucosa presenta grandes
pliegues primarios y pliegues secundarios menores, muy numerosos en el pabelln, los que
van disminuyendo en direccin al cuerno. Su epitelio de clulas cilndricas, asienta sobre
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
una base de tejido conectivo ricamente vascularizado, y est constituido por las clulas
ciliadas ms numerosas a nivel del pabelln cuyos cilios se mueven en direccin contraria
al ovario, lo que crea en el interior del oviducto una corriente hacia el tero [38], y por las
clulas glandulares.
Las ciliadas tienen importancia en la denudacin del vulo antes de la fecundacin e
intervienen en la migracin del cigoto. Las glandulares en la vaca, secretan despus que el
cigoto ha emigrado a travs de la trompa, posiblemente esta secrecin al fluir hacia el
tero, le proporcione elementos nutritivos necesarios para su segmentacin y desarrollo,
hasta su fijacin en la mucosa uterina [14].
Las clulas epiteliales de los oviductos de la vaca, de 26-28 de alto durante la fase
lutenica, se congestionan e hipertrofian en la fase folicular, y llegan a 47 u en el celo [14].
El edema y la presencia de grnulos, alcanzan su punto mximo desde el primer da del
pro-estro, hasta el segundo da, mientras que las clulas ciliadas conservan su actividad en
todo el ciclo [39]. La aparicin y la desaparicin de leucocitos siguen un curso paralelo a la
presentacin de grnulos en las clulas epiteliales.
La muscular, constituida por fibras lisas en el pabelln, se encuentra bien desarrollada en la
ampolla y an ms a nivel de la conjuncin tero-tubrica. Se hallan dispuestas, las
internas en direccin circular y las externas en sentido longitudinal, lo que le permite dar al
rgano un movimiento peristltico, similar al intestinal.
El papel desempeado por las contracciones tero-tubricas, en el transporte de los
espermatozoides a travs del tracto genital, fue demostrado desde 1952 por Van Dermark y
Hays [40], quienes mediante la excitacin de la vagina o del cuello uterino, por el coito o
por la inseminacin artificial, desencadenaron una serie de contracciones uterinas y
tubricas, que tuvieron como resultado el ascenso de los espermatozoides, hasta las partes
altas del oviducto. Estas contracciones parecen estar ligadas a una descarga refleja de
oxitocina [25]. El descenso del cigoto a travs del oviducto depende de los estregones y de
la progesterona, y es favorecido por esta.
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
Ciclo uterino
El tero est constituido por una mucosa, una lmina muscular y una serosa que lo cubre
completamente.
La mucosa uterina o endometrio posee un epitelio secretor cilindrito pseudos- estratificado.
La lmina propia contiene las glndulas uterinas, que son tubulosas, ramificadas y ms o
menos tortuosas, especialmente hacia su extremo. Las glndulas poseen una membrana
propia laminar y estn revestidas de un epitelio cilndrico simple. Son mucho ms anchas
en su superficie que hacia la profundidad y su extremo libre siempre se sita entre los
espacios intercarunculares. La mucosa uterina carece de submucosa, las glndulas llegan
hasta la muscular, y en ocasiones internan en estas su extremo ciego [41].
El endometrio de la vaca presenta de 80-120 carnculas de 1-2 mm de altura, dispuestas en
cuatro lneas longitudinales. En su base son muy abundantes los vasos capilares. Durante la
gestacin estas carnculas se hipertrofian, algunas alcanzan de 8-10 cm de largo, y
presentan una estructura esponjosa con numerosas criptas en su superficie, en la que
penetran y se fijan las vellosidades coriales de los cotiledones, para constituir los
placentomas.
En el ciclo estral, se producen modificaciones en el tero, influidas por las hormonas
gonadales, las que son ms notables en el endometrio. En la fase folicular, en los das del
18-21 y 22 del ciclo estral, con predominio de estrgenos, las modificaciones se
manifiestan por un aumento de la vascularizacin, que se intensifica a medida que crece
bajo la accin del estradiol y el edema que se inicia en las profundidades del estroma y
avanza hacia la superficie.
Con esto aumenta el espesor del estroma, lo que provoca la disminucin de la tortuosidad
de las glndulas uterinas; ya en los das 1 y 2 del ciclo, el epitelio alcanza su secrecin
mxima, de tipo holocrino.
En la vaca hay acumulo de glucgeno en la superficie epitelial [14], a diferencia de la
mujer, en la que este es muy abundante en la fase progesternica, y aparece una infiltracin
de polimorfo nucleares que alcanza su mxima intensidad los das 3 y 4 post-estro. Se ha
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
encontrado mayor concentracin de leucocitos en la mucosa y el lumen del tero, en
respuesta a la inoculacin de bacterias, durante el estro, que en la fase luteal. Los
plasmocitos son ms abundantes a nivel del tero de vacas viejas, que en las jvenes o en
las novillas.
En la fase lutenica progestativa, con predominio de progesterona, el estroma sometido a
una deshidratacin fuerte, tiene inclinacin a la hemorragia, las glndulas uterinas crecen
muy rpido, tanto en longitud como en dimetro, se ramifican y adquieren un curso
tortuoso, su secrecin es de tipo apocrino y merocrino. En la vaca la fase lutenica est
caracterizada por la presencia de fosfatasa alcalina.
Al producirse la ovulacin hay una cada de los niveles de estrgenos, y los vasos
sanguneos congestionados dan muestras de fragilidad, extravasndose pequeas
cantidades de sangre en el tero, la que fluye por el aparato reproductivo y con frecuencia
impregna la cola y la regin perineal de la vaca. Un crecida nmero de novillas y
aproximadamente la mitad de las vacas, muestran signos visibles de hemorragia entre las
48 y 96 h del comienzo del celo.
La muscular uterina o miometrio, es la ms gruesa de las tres capas que componen el tero.
Est dividida por un estrato vascular, que posee los importantes vasos sanguneos que
irrigan al tero, y por fibras nerviosas, en una capa interna de fibras circulares y una
externa de fibras longitudinales. A veces en la vaca el estrato vascular est situado dentro
de la mitad externa de la capa muscular de fibras circulares [41]. Las musculares varan de
tamao en el transcurso del ciclo y aumentan su longitud en la fase folicular, en la que
alcanza de 110-120 y disminuyen en la lutenica a una media de 70-80
La capacidad contrctil del tero est supeditada al equilibrio estrgeno progesterona [14].
Su actividad espontnea se incrementa en el estro. Con ayuda de las contracciones uterinas,
los espermatozoides pueden ser transportados a los oviductos, a los pocos minutos de la
deposicin del semen. Este rpido transporte de los espermatozoides ha sido demostrado
en la vaca [42] y [43].
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
Durante la gestacin, las fibras musculares lisas del miometrio se distienden enormemente
y son sensibles a las hormonas que causan las contracciones uterinas en el momento del
parto. El desarrollo muscular se inicia en la extremidad de los cuernos uterinos y progresa
hacia el cuerpo del rgano. La regresin sigue el mismo orden.
Ciclo cervical
El cuello uterino constituye un fuerte esfnter muscular, tubular, que mientras permanece
cerrado es capaz de proteger al tero de la invasin de grmenes dainos. Sus paredes son
gruesas, duras y ms rgidas que las del tero o la vagina, especialmente en las unparas.
Presenta de 3-5 pliegues transversales o anillos, de consistencia casi cartilaginosa y es
difcil o imposible dilatarlo manualmente, pero en el estro se produce una ligera dilatacin
para dejar pasar a los espermatozoides y en el parto se dilata en su totalidad, quedando
borrado para dar paso al feto.
La mucosa cervical carece de glndulas, las que nicamente existen en los carnvoros, pero
en la vaca posee altos pliegues longitudinales, provistos de plieguecitos secundarios,
limitados por surcos longitudinales profundos anfractuosos, que se subdividen en surcos
menores de aspecto glandular [41] que aumentan en gran medida su superficie. Su epitelio
est provisto de numerosas clulas mucgenas, que segregan gran cantidad de mucus,
secrecin cuyo volumen, naturaleza y composicin, vara de acuerdo con el perodo del
ciclo estral.
En la fase estrognica (la de mayor actividad), hay congestin sangunea y edematizacin,
sus clulas epiteliales cilndricas, son muy altas, con numerosas mitosis, que le dan al
epitelio aspecto de estratificacin. El da 17 del ciclo en los inicios de la etapa se puede
observar la preparacin del epitelio para la secrecin de mucus. La mxima replecin de
las clulas mucgenas se alcanza al inicio del estro, cuando se rompen por su pice y
descargan su contenido mucoso en la luz cervical.
En el inicio del celo la cantidad de moco estral es muy abundante y se acumula en el fondo
de la vagina. Al progresar el estro y ser mayor el volumen de dicha secrecin, esta sale al
exterior, clara, transparente, viscosa, con abundantes restos celulares, y muy filante, lo que
le permite colgar de la vulva. El moco cervical tiene un pH alto, y gran capacidad de
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
cristalizacin (arborizacin) en el curso de su desecacin; esta cristalizacin forma cuadros
tpicos parecidos a las hojas de helecho [17], que se pueden observar al microscopio con el
menor aumento.
En esta fase de abundante secrecin algunas clulas pierden su ncleo y sufren
degeneracin [44]. La gran actividad que desarrollan las clulas mucgenas, y los
caracteres del moco segregado, parecen depender del aumento de la vascularizacin del
estroma que lo preceda.
En la fase progesternica, decrece la congestin hasta llegar a desaparecer, o ser muy pobre
alrededor de 6 das despus de terminado el celo. En sus inicios el moco se hace
opalescente, diminuye, se condensa y pierde su capacidad de cristalizacin. Tras la
abundante secrecin ocurrida en el estro, la mayora de las clulas mucgenas conservan su
ncleo, regeneran su citoplasma y recobran su capacidad funcional [44].
En los animales gestantes el mucus cervical forma un tapn mucoso que sella el tero
durante la gestacin, su ruptura en el transcurso de esta provoca aborto o maceracin fetal.
Ciclo vaginal
La vagina, rgano de la cpula, de pared delgada, pero resistente y muy elstica, tanto que
su dilatacin solo se ve limitada por la pared de la cavidad pelviana [45], consta de
mucosa, una muscular mal definida, la adventicia y la serosa.
La mucosa presenta pliegues longitudinales y carece de glndulas, en su porcin anterior
forma pliegues transversales. El mucus que normalmente se encuentra en la mucosa
vaginal y que aumenta de manera considerable en las hembras en celo, procede
principalmente del crvix, desde donde fluye al lumen vaginal [38]. Su epitelio es
cilndrico y aumenta su grosor en el tercio craneal y se convierte en estratificado en la
porcin caudal. En la vaca sus clulas superficiales pueden cornificarse en cierto grado. La
lmina propia en esta especie carece de cuerpo papilar o lo tiene mal definido; su tejido
conjuntivo elstico, rico en clulas en la regin infraepitelial contiene ndulos linfticos
[41].
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
En el epitelio vaginal ocurren tambin cambios cclicos. Muy delgado en el diestro, con
clulas cuboidales en su superficie y leucocitos que migran continuamente a travs del
epitelio al lumen. En el pro-estro y principios del estro hay un crecimiento marcado e
incremento del grueso del epitelio, producido por la divisin de las clulas de su capa
basal, las que tienden a hipertrofiarse y hacerse poligonales segn se extienden hacia la
superficie. Estas clulas, cerca del lumen son aplastadas, pero retienen su ncleo.
En la vaca hay una extensa formacin de clulas, en forma de copa, en la regin craneal de
la vagina, que segregan mucus. Durante el pro-estro y principios del estro se incrementa la
cantidad de mucus, y en el metestro su produccin decrece, se hace ms denso y toma un
color blancuzco 0amarillento, de apariencia opaca, a causa de la presencia de gran cantidad
de epitelio descamado [46].
En frotis realizados en la vagina de la vaca, en el perodo estral, se han encontrado algunas
clulas cornificadas, grandes clulas epiteliales y numerosos leucocitos, los que
disminuyen en el metestro, mientras que las clulas epiteliales predominan hasta el da 8
del ciclo. Las clulas cornificadas aumentan mucho a partir del da 9 y son abundantes
hasta el da 16 o 17. Estas modificaciones no son lo suficientemente pronunciadas, para
permitir el diagnstico fcil y eficaz del estadio del ciclo estral [14], como sucede en los
roedores.
La muscular posee una capa interna de fibras circulares, y otra externa ms delgada, de
fibras longitudinales. En la cerda y en la perra existe una delgada capa de las longitudinales
por dentro de la capa de las circulares [41].
La adventicia, de naturaleza conjuntiva, contiene grandes vasos, nervios y ganglios.
Ciclo vestibular
El vestbulo vaginal est limitado cranealmente por el anillo himenial, en el que se puede
ver el rudimento de himen, formado por un repliegue de la mucosa vaginal [8]. Constituye
un segmento comn a los tractos genital y urinario, que toma origen del seno urogenital.
Cuenta con mucosa, muscular lisa cubierta por una estriada y una adventicia que fija el
vestbulo a los tejidos de la pelvis, falta la serosa que cubre el resto del tracto genital.
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
La mucosa est cubierta por una lmina epitelial estratificada que a menudo es atravesada
por un nmero crecido de leucocitos [41]. Alrededor de la mucosa existe una fuerte
musculatura lisa, que cuenta con dos capas, una interna circular y otra externa cuyas fibras
son longitudinales. Cubriendo a las fibras lisas hay una capa muscular de fibras estriadas,
que constituyen los msculos constrictores del vestbulo y de la vulva. Estos msculos, con
su fuerte contraccin contribuyen al cierre del canal vaginal y al aislamiento de los rganos
genitales internos, con el medio exterior [8] y su dilatacin tiene importancia en el
momento del parto.
El tegumento de los labios vulvares posee una extructura similar a la de la piel. Tiene
abundantes glndulas sebceas y tubulosas. La superficie externa de los labios vulvares
est tapizada por piel fina, muy delgada, posee fibras elsticas y numerosas glndulas
sebceas. Su mucosa est cubierta por un epitelio estratificado. Los labios tienen fibras
musculares lisas y, adems, fibras musculares estriadas, dispuestas circularmente alrededor
de la hendidura vulvar (constrictor cunni) [41].
Al igual que sucede con el resto del aparato genital, este segmento se ve influido por las
diferentes fases del ciclo estral. En el pro-estro la vulva se congestiona, la mucosa se
aprecia edematosa, hipermica, brillosa, de color rosado, al llegar el estro, estas
manifestaciones ganan en intensidad llegando la mucosa a tomar un color rojo claro. En el
metestro desaparece la edematizacin y la hiperemia, la mucosa toma un color rosado que
se torna plido en el diestro.
Variaciones en el pH
El pH vaginal por lo general se encuentra entre 6 y 7 y es ms elevado en el estro que en el
diestro. El crvix es ms cido que la vagina.
Las determinaciones in vivo, son aproximadamente una unidad de pH mas bajas, que las
efectuadas in vitro, sobre el mismo moco.
Resulta interesante, que al contrario de lo que se puede suponer, las infecciones del aparato
genital de la hembra, elevan su pH en vez de reducirlo. El aumento interesa a la zona
infectada y a las partes comprendidas entre esta y el exterior.
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
REFERENCIAS BIBLIOGRFICAS
1. HOPKINS, S.M.: Patrones reproductivos del ganado bovino. En: Endocrinologa
veterinaria y reproduccin. (Ed.) L.E. Me Dnalo, Nueva Editorial Interamericana Me
Graw-Hill, Mxico D.F-, 1991.
2. PINEDA, M.H.: Sistema reproductor de la hembra.En: Endocrinologa veterinaria y
reproduccin. (Ed.) L.E. Me Donald, Nueva Editorial Interamericana Me Graw-Hill,
Mxico D.F, 1991.
3. JAINUDEEN, M.R. y E.S.E. HAFEZ: Ganado bovino y bfalo de agua. En:
Reproduccin e inseminacin artificial en animales. (Ed.) E.S.E. Hafez, 1989.
4. GARVERICK, H.A. and M.F. SMITH: Female reproductive physiology and
endocrinology of cattle. Veternary Clinics of North Amrica Food Animal Practico, 9
(2): 223,1993.
5. HAFEZ, E.S.E.: Reproduction in farm animals. 4th ed. Lea and Febiger, Philadelphia,
1980.
6. SORENSEN, A.M.; W.H. HOUGHT; D.T. ARMSTRONG et al.: Causes and
prevention of reproductiva failures in dairy cattle I. The influence of underfeeding and
overfeeding on growth and development of Holstein heifers. Come// Unk Agrie. Exp.
Sta. Bull., 936,1959.
7. BRITO, R.; N. PERN y G. MARTNEZ: Estudio de la conducta reproductiva de la
raza Ceb. Informe de tema de investigacin, Ministerio de la Agricultura, La Habana,
1980.
8. HOLY, L: Biologa de la reproduccin bovina. Edicin Revolucionaria, Instituto
Cubano del Libro, La Habana,1987.
9. LAM ROMERO, F.: Contribucin al estudio de los principales indicadores sanguneos
del metabolismo proteico y energtico de hembras bovinas en desarrollo. Tesis para
optar por el grado de Doctor en Ciencias Veterinarias, La Habana, 1986.
10. THOMPSON, J.K.; D.C. Me DNALO and R.W. WARREN: Mltiple blood analysis
of dairy cows as a management aid. The Scoland College of Agriculture (january),
1978.
11. FOREST, M.G. and M.C. LEVASSEUH Puberty. En: Reproduction in mammals and
man. (Eds.) C. Thibault; M.C. Levasseur and R.H.F. Hunter, Ellipses, Pars, 1993.
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
12. GONZLEZ-PADILLA, E.; J.N. WILJBANK and G.D. NISWENDER: Puberty in
beef heifers I. The interrelationship betwen pituitary, hypotalamic and ovaran
hormones. J. Anim. Sci., 40(6): 1091,1975.
13. Mitchell, D. and W.R. Alien: Observations on reproductive perfomance in the yearling
mare. J. Reprod. Frtil., 23: 531- 536,1975.
14. DERIVAUX, J.: Reproduccin de los animales domsticos. Editorial Acribia,
Zaragoza,1976.
15. LEVAUSSER, M.C. and C. THIBAULT: Reproductive life cycle, En: Reproduction
in farm animals. (Ed.) E.S.E. Hafez, Lea and Febiger, Philadelphia, 1980.
16. Me DNALO, L.E.: Endocrinologa veterinaria y reproduccin. Nueva Editorial
Interamericana Me Graw-Hill, Mxico, D.F., 1991.
17. MATTON, R; V. ADELAKOUN; Y. COUTURE and J.J. DUFOUR: Growth and
replacement of the bovine ovaran follicles during the estrous cycle., J. Anim. Sci.
52:813-820,1981.
18. CUPPS, RT; L.L. ANDERSON and H.H. COL: The estrous cycle. En:
Reproduction in domestic animis, (Eds.) H.H. Col and RT. Cupps, Academic Press,
New York and London, 1969.
19. CONCANNON, RW. J.R Me CANN y M. TEMPLE: Biology and endcrinology of
ovulation, pregnancy and parturition in the dog. J. FSeprod. Feri. Suppl., 39,325,1989.
20. CHAFFAUX, S,: Reproduction of the cat and the dog. En: Reproduction in
Mammals and Man. (Eds.) C. Thibault; M.C. Levasseur y R.H.F. Hunter, Ellipses,
Paris, 1993.
21. TSAFRIRI, A.; N. DEBEL and S. BAR-AMI; The role of oocyte maturation Inhibitor
In follicular regulation of oocyte maturation. J. feprod. Frtil., 64: 541, 1982.
22. ROBERTS, S.J.: Veterinary obstetrics and genital diseases. Edwards Brothers, Inc. Ann
Arbor, Michigan, 1971.
23. ERICKSON, B.H.; Development and radio response of the prenatal bovine ovary. J.
Reprod. frtil. 11: 97,1966.
24. PINEDA. M.H.: La biologa del sexo. En: Endocrinologa veterinaria y
reproduccin. (Ed.) L.E, Me Donald, Nueva Editorial Interamericana Me Graw-Hill,
Mxico, D.F., 1991.
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
25. AREY, L.B.: Anatoma del desarrollo. Edicin Revolucionara, Instituto Cubano del
Libro, La Habana.1968.
26. RAJAKOSKI, E.: The ovarian follicular system in sexuallly mature heifers with
special reference to seasonal, cyclical and lef right variations. Acta Endocrinolgica,
suppl., 52 (L11) vol XXXII, 1960.
27. Variaciones estacionales y cclicas del sistema folicular ovrico en el ganado.
Procceding of the IV International Congress on Animal Reproduction, The hague, vol.
II. 1961.
28. SIRIOS, J. y J.E. FORTUNE: Ovarian follicular dynamics during the estrous cycle in
heifers monitored by real-time ultrasonograpny. Biol. Reprod. 39:308-317,1988.
29. PIERSON, R.A. and O.J. GINTHER: Ultrasound imaging of the ovaries and uterus in
cattle. Theriogenology, 29: 21-37,1988.
30. GRENSWALD, G.S. and RF. TERRANOVA: Follicular selection and its control.
En: Thephysiologyofreproduton, (Eds.) E. Knobil; J. Neill et al. Raven Press, Ltd.,
New York, 1988.
31. BRITT, J.A.: Currents concepts of folliculogenesis and endocrinology, Embryo.
Transfer., 3(4): 1-3,1988.
32. DRIANCOURT, M.A.; A. GOUGEON; D. ROYERE y C. THIBAULT; Ovarian
function. En: Reproduction in mammals and man. (Eds.) C. Thibault; M.C. Levasseur
and R.H.F. Hunter. Ellipses, Paris, 1993.
33. STABENFELDT, G.H.; J.R HUGHES and J.W. EVANS: Ovaran activity during the
estrous cycle of the mare. Endocrinology, 90:1379-1384, 1972.
34. HAFEZ, E.S.E.: Reproduccin e inseminacin artificial en animales. Nueva Editorial
Interamericana Me Graw-Hill, Mxico D.F., 1989.
35. OSMAN, R Y J. DULLART: "Intraovarian relase of eggs in the rat after indomethacin
treatment of proestrus". J. Reprod. Frtil. 47:101,1976.
36. MARIN, G.B.; V.R. SMITH; T.E. WILLEY and G.R. BARRET: The effects of
sterile copulation on the time of ovulation in dairy heifers. J. Dairy ScL, 33:885,1950.
37. PETERS, A.R.: Hormonal control of the bovine oestrous cycle I. The natural cycle.
Br. Vet. J., 141: 564-575, 1985.
38. NALBANDOV, A.Y: Fisiologa de la reproduccin. Ed. Acribia, Zaragoza, 1969.
39. LOMBARD, L; B.B. MORGAN and S.H. Me Nun: The morphology of the oviduct of
virgin heifers in relations to the estrous cycle. J. Morph., 86:1-24,1950.
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
40. VAN DENMARK, N.L. and R.L. HAYS: Uterine motility responses to mating. Am.
J. PhysiologyMQ: 518-521,1952.
41. TRAUTMANN, A. y U. FIEBIGER: Histologa y anatoma microscpica comparada
de los animales domsticos. Editorial Labor, S. A., Barcelona, 1942.
42. BIRGITTA, L.: Distribution of spermatozoa in the bovine genital tract after artificial
insemination. Thesis, Department of Obstetrics and Gynecology. Faculty of Veterinary
Medicine, Swedish University Agricultural Sciences, Uppsala, Sweden, 1988.
43. VAN DENMARK, N.L. and A.N. MOELLER: Speed of spermatozoan transport in
reproductive tract of estrous cow. Am. J. Physiology, 165:674-679,1951.
44. HERRICK, J.B.: The citological changes in the cervical mucosa of the cow (Bos
taurus) throughout the estrous cycle. Am. J. Vet. Res., 12:246-281,1951.
45. SALISBURY, G.W. y N.L. VAN DEMARK: Fisiologa de la reproduccin e
inseminacin artificial de los bovidos. Edicin Revolucionaria, Instituto Cubano del
Libro, La Habana,1969.
46. COL, H.H. y RT CUPPS: Reproduction in Domestic Animals. Academic Press, New
York and London, 1969.
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
3. REGULACIN NEUROENDOCR1NA DEL CICLO ESTRAL
Hace cincuenta aos se pensaba que la homeostasis hormonal era mantenida por un
mecanismo puramente endocrino con el que se explicaba el comportamiento del ciclo
estral.
Hoy da se sabe que el sistema de control que regula los procesos endocrinos est
constituido por un mecanismo hormonal y un sistema regulador nervioso, los que
funcionan en estrecha relacin.
Estos dos sistemas reguladores aparentemente son muy distintos, ya que en el endocrino la
informacin se transmite por la sangre (recurdese la definicin de hormona) y en el
vegetativo mediante impulsos nerviosos, que permiten respuestas ms rpidas. Sin
embargo, las diferencias no son siempre tan claras porque las terminaciones nerviosas
liberan transmisores qumicos, colinrgicos y adrenrgicos (acetilcolina y noradrenalina),
los que, en ocasiones, circulan por la va hemtica y pueden ser considerados, por tanto,
como hormonas. Por otra parte, la clsica definicin de hormona requiere en la actualidad
tambin revisin.
Ambos sistemas, vegetativo y endocrino, tienen un centro integrador comn, en el que se
funden sus actividades, el hipotlamo, que, adems, de realizar sus funciones nerviosas es
el lugar de sntesis de varias hormonas polipeptdicas. De aqu que el sistema nervioso
vegetativo y las diversas glndulas endocrinas representen, en el ms amplio sentido de la
palabra, segn Rasmussen, un solo sistema neuroendocrino que ha evolucionado para
integrar y coordinar las actividades metablicas del organismo.
Conceptos bsicos
Feedback. El concepto general ms importante es el de control por retroaccin
(feedback), especialmente el de retroaccin negativa, aunque la positiva puede ser
importante en determinados casos.
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
Pulso. Un pulso puede ser definido como un episodio de liberacin hormonal, corto pero
intenso. Los dos principales parmetros que definen las caractersticas de la liberacin
pulstil son: la frecuencia de ocurrencia y la amplitud de los pulsos.
Tasa basal. Al parecer existe una tasa basal de secrecin de todas las hormonas, y cuando
las circunstancias lo requieren, las tasas de secrecin aumentan notablemente.
Los rganos diana pueden funcionar a un nivel mnimo aun en ausencia de la hormona. De
ese modo la naturaleza ha prevenido un nivel basal de funcin que puede ser suficiente
para mantener vida en su ausencia, por ejemplo, todos los tejidos son capaces de usar
carbohidratos a un nivel mnimo incluso en la ausencia de insulina.
Vida media. La vida media que disfruta una determinada hormona en el plasma puede
variar entre unos minutos y varios das. Tambin la respuesta fisiolgica apreciable de un
tejido particular puede ser inmediata o presentarse despus de varias horas, as como una
misma hormona puede suscitar una respuesta inmediata en un tejido y una respuesta tarda
en otro.
Receptores. Todas las clulas estn expuestas a las hormonas, pero solo algunas de ellas
pueden responder por poseer un sistema receptor altamente especfico, por lo que son
llamadas clulas diana. Estos receptores en el caso de las hormonas peptdeas, que tienen
molculas grandes y por ello no pueden pasar a travs de las membranas plasmticas, se
encuentran situados en la superficie de las clulas. Las hormonas esteroides y tiroideas,
que por tener molculas pequeas pueden atravesar dicha membrana celular, tienen los
receptores en el citoplasma para las hormonas esteroides y en la cromatina nuclear para las
tiroideas.
Sistemas de control endocrino. Existen distintos tipos de sistemas de control endocrino.
El ms simple parece ser un sistema en el que la hormona liberada de la glndula endocrina
acta sobre la clula sensible, la que libera una sustancia que mediante retroaccin
negativa disminuye la tasa de liberacin de la hormona (Figura 3.1). Un rasgo bsico de
este tipo de sistema es la falta de control hipotalmico o hipofisario directo.
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
FIGURA 3.1 Sistema simple de control endocrino en el que solo intervienen la glndula endocrina y la
clula efectora.
En un sistema ms complejo la actividad de la glndula endocrina esta bajo el control del
hipotlamo, mediante la ordenacin jerrquica, hipotlamo-glndula endocrina-clula
electora. La seal aferente inicial del hipotlamo puede ser nerviosa u hormonal (factor de
liberacin), esta seal va dirigida a la glndula endocrina, la que segrega la hormona que
va a actuar sobre las clulas eferentes. Estas provocan un cambio en un componente
plasmtico el que regula la actividad hipotalmica. La caracterstica que diferencia este
tipo de sistema es que el control por retroaccin no se ejerce de manera directa sobre la
glndula, sino sobre la funcin hipotalmica, que a su vez controla la funcin glandular
(Figura 3.2).
FIGURA 3.2 Sistema complejo en el que interviene el hipotlamo. El control por retroaccin lo ejerce la
clula efectora.
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
En el mximo orden de complejidad de estos sistemas, la actividad del rgano elector
endocrino final se regula por la hipfisis anterior, cuya actividad es controlada por el
hipotlamo. Su ordenacin jerrquica es: hipotlamo-hpfisis anterior-glndula endocrinaclula efectora. En este caso el hipotlamo enva una seal, factor de liberacin al lbulo
anterior de la hipfisis, este libera una hormona que acta sobre la glndula endocrina, la
que elabora las hormonas finales que inciden sobre las clulas efectoras y van a ejercer el
control retroactivo del hipotlamo (Figura 3.3).
FIGURA 3.3 Sistema de mxima complejidad, en el que intervienen el hipotlamo y la hipfisis. El control
hipotalmico se ejerce mediante la glndula endocrina.
Este complejo sistema est caracterizado por dos rasgos importantes, el primero, que el
elector del mecanismo de retroaccin es el producto endocrino final, y no alguna sustancia
que resulta de su accin sobre las clulas sensibles, y el segundo, que es el hipotlamo el
asiento del control por retroaccin.
A pesar de la complejidad de este sistema, la actividad cclica de las gnadas femeninas y
la funcin uterina requieren una complicacin mayor para su regulacin y es por ello que
las hormonas finales estradiol y progesterona, mediante actividades de retroaccin negativa
y positiva, coordinan la secrecin cclica de los factores hipotalmicos e hipofisiarios.
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
Los sistemas descritos son cerrados, no obstante su equilibrio final puede ser modificado
por la accin del medio exterior sobre sus componentes, hipotlamo, hipfisis, glndula
endocrina y clulas efectoras.
Mecanismo de accin de las hormonas proteicas
Las hormonas proteicas, que se unen a un receptor en la membrana celular, estimulan a la
enzima adenilciclasa, para convertir ATP al nucletido adenosn monofosfato (3,5 AMP o
AMPc).
El AMPc -segundo mensajero, por propagar a travs de la clula el efecto de la hormona,
primer mensajero- provee energa para la sntesis proteica, y la protena inducida estimula
a la mitocondria a producir otra hormona, tercer mensajero [1] (Figura 3.4).
FIGURA 3.4 Modo de accin de una hormona proteica [1]
Mecanismo de accin de las hormonas esferoides
El mecanismo de accin de las hormonas esferoides es diferente al de las proteicas. Las
primeras por poseer molculas pequeas se difunden libremente en muchas clulas del
cuerpo, ya que la membrana plasmtica no constituye para ellas una barrera. Pero solo las
clulas diana poseen los receptores especficos, a los que se puede unir la hormona, Estos
receptores para las hormonas esferoides se encuentran en el citoplasma.
Cuando una hormona esferoide penetra en la clula diana que posee su receptor especifico,
se une a este y forma un complejo hormona-receptor que penetra en el ncleo celular
donde hay un punto del cromosoma que lo acepta y que contiene los genes (ADN) de la
clula. En este sitio efector el complejo hormona-receptor causa la sntesis de un mensajero
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
ARN que depende de un gen determinado. Este mensajero entonces migra al retculo
endoplsmico donde ordena la sntesis de una nueva protena (Figura 3.5).
Transporte de las hormonas
Las hormonas proteicas, usualmente se conservan en las glndulas en que son producidas,
hasta que se necesitan, momento en que son secretadas en los diferentes capilares
sanguneos. Las hormonas esferoides no se almacenan sino se liberan tan pronto como son
producidas [1].
Por lo general las hormonas esferoides se unen a globulinas especficas que contiene el
plasma, lo que incrementa su solubilidad en el medio acuoso de la sangre y restringe su
difusin a travs de los tejidos, al mismo tiempo que prolonga su accin al protegerla
contra la degradacin y la eliminacin.
La forma unida de una hormona es inactiva a los niveles intracelulares, pero est en
equilibrio con una pequea cantidad de la forma libre, y as es tomada por las clulas
diana.
FIGURA 3.5 Modo de accin de una hormona esteroide [1]
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
Hipotlamo
Los primeros en sugerir la intervencin del hipotlamo en e) control de la funcin de la
glndula hipfisis fueron Green y Harris [2]. Las clulas nerviosas que se encuentran en el
hipotlamo son idnticas a las del cerebro y tienen la misma capacidad para recibir y
transmitir un impulso nervioso. Pero se diferencian de las clulas nerviosas en que no
inervan a ninguna clula y en que terminan en fondos de saco de los vasos sanguneos de la
neurohipfisis; adems, algunas neuronas hipotalmicas contienen grnulos de origen
secretorio, de aqu la denominacin de neurosecretoras que se le da a estas clulas y el de
neurosecrecin para el producto que ellas contienen [3].
El hipotlamo produce sustancias polipeptdicas que estimulan o inhiben la liberacin de
las seis hormonas adenohipofisarias, los factores de liberacin o releasing hormn (RH).
Estas son transferidas a la hipfisis anterior mediante la va del sistema portadienceflico,
lo que permite que con una pequea cantidad de RH se pueda elevar su nivel en la
hipfisis, mientras que su nivel sistmico es bajo [1].
Factores de liberacin
Hoy da se sabe que los factores liberadores comprenden una familia de polipptidos de
bajo peso molecular. El primero en ser descubierto fue el factor de liberacin de
corticotropina (CRF), en 1955 y cinco aos ms tarde el releasing hormn (que ms inters
tiene para nosotros) de la hormona luteinizante (LH), denominado LHRH o LRH, es un
decapptido que se considera acta como liberador de la LH y de la FSH, por lo cual en la
actualidad se le conoce como gonadotropin releasing hormn (GnRH).
Algunas de estas sustancias hipotalmicas actan no como liberadoras sino como
inhibidoras, y lo hacen inhibiendo la liberacin de su hormona correspondiente, la ms
conocida es el factor de inhibicin de la prolactina de los mamferos (PIF).
Es interesante sealar que los extractos hipotalmicos portadores de estos factores
liberadores, tambin, contienen cantidades apreciables de hormonas fisiolgicamente
similares, y tal vez idnticas a las propias hormonas hipofisarias. Se ha demostrado la
presencia de LH, FSH, TSH y ACTH en extractos de intensa actividad liberadora. No se
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
sabe cul puede ser el significado fisiolgico de esta coexistencia de factores liberadores y
hormonas en el hipotlamo y en la eminencia mediana [4].
Despus de la administracin de un factor de liberacin hay respuesta hipofisaria en
cuestin de minutos y su vida media en la circulacin tambin es corta, igualmente de
algunos minutos. Parece que son qumicamente idnticos en diferentes especies animales,
esta falta de especificidad simplifica su aplicacin en los animales.
Para tener una idea de la potencia de estas sustancias se puede decir que se necesitan ms
de 3 kg de hipotlamo de cerdas para producir 1 mg de GnRH, pero la dosis mnima
efectiva para una rata puede ser tan baja como 5 pg [5].
Un aspecto interesante de la fisiologa de los RH es el descubrimiento de que el nivel de
GnRH en la leche de las vacas, las mujeres y/o las ratas excede al nivel en el suero, lo cual
implica su concentracin en la glndula mamaria o un sitio extrahipotalmico de
produccin de GnRH. La GnRH en leche puede afectar las gnadas de neonatos y otros
[6].
GONADOTROPIN RELEASING HORMN (GnRH)
La GnRH es un decapptido de peso molecular 1 182 Dalton, compuesto de los
aminocidos:
PYRO-GLV-HIS-TRP-SER-GLY-LEU-ARG-PRO-GLY-NH.
Esta
composicin es idntica en todos los mamferos, incluido el hombre.
El principal rgano efector de esta hormona es la adenohipfisis, en la que causa la
liberacin de las principales hormonas gonadotrpicas, la luteinizante (LH) y la folculo
estimulante (FSH).
La GnRH estimula una rpida secrecin de LH y secundariamente su biosntesis en orden
de restaurar su existencia, por consiguiente, es la moduladora esencial de la secrecin de
LH, y tanto los factores internos [feedback gonadal] o externos (fotoperodo, feromonas,
etc.) ejercen su principal accin a travs de la modulacin de la secrecin de GnRH por el
hipotlamo [7].
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
Por contraste la GnRH parece ser indispensable para el mantenimiento de un nivel
suficiente de sntesis de FSH, la que es modulada por diferentes factores gonadales [7].
La secrecin de GnRH es un fenmeno biolgico caracterizado por su naturaleza pulstil.
Cada pulso de GnRH es el producto de la adicin de pequeas cantidades de la hormona
liberada por diversas neuronas secretoras de GnRH. La naturaleza pulstil de su secrecin
es el resultado de un ritmo interno de 15 - 30 min (dado por un marcapasos) y de la
sincronizacin entre las neuronas secretoras [5].
La GnRH in vitro estimula la secrecin por clulas de la hipfisis de las gonadotropinas
hipofisarias FSH y LH, pero como estas dos hormonas no son secretadas in vivo
sincrnicamente, est claro que existe una regulacin independiente para cada una de ellas
[7]. Muchas observaciones soportan la existencia de una hormona liberadora de FSH
(FSHRH) diferente de la GnRH.
En 1971 se logr la sntesis de la GnRH, dos gonadorelinas originales tienen en los
EE.UU. las marcas comerciales Cystorelin y Factrel. Algunas modificaciones en la
estructura qumica de la molcula original de GnRH dan lugar a potentes anlogos. Su
potencia vara con la secuencia de sus aminocidos, la sustitucin de las posiciones que
estos ocupan y el aminocido especfico cuya posicin es modificada. En la actualidad
existen diversos anlogos comerciales que se utilizan satisfactoriamente en la prctica,
entre ellos Buserelin (alemn), Fertagyl (holands), Ferterelin (japons) y Deslorelin
(australiano). Ferterelin es de 4-10 veces ms potente que Gonadorelin, y Buserelin y
Deslorelin son 50 veces ms potentes.
Dosis de 100 g de GnRH sinttica producen en la vaca una respuesta equivalente a la
descarga de LH que precede a la ovulacin. Mientras que la LH liberada aumenta de forma
lineal hasta una dosis de 1 500 g de GnRH, la descarga de FSH es creciente hasta 500 g
de GnRH, dosis a la cual Se obtiene la respuesta mxima (Figura 3.6).
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
FIGURA 3.6 Relacin dosis respuesta para LH y FSH la inyeccin IM de GnRH [8]
La aplicacin intramuscular de 200 g de GnRH sinttica provoca en 15 min la liberacin
de FSH y de LH, con la produccin de un pico a los 120 min. El regreso a los niveles
bsales se origina entre 300 y 360 min [8] y [9]. La aplicacin por va intravenosa obtiene
una respuesta ms rpida, pero la cantidad de gonadotropina liberada alcanza niveles
similares.
Neuromediadores en el hipotlamo
Neuromediador. Es un trmino general que se aplica a todas las sustancias sintetizadas,
almacenadas y secretadas por neuronas como respuesta a mensajes neurales. De acuerdo
con su modo de accin los neuromediadores son llamados neurotransmisores,
neurohormonas o neuromoduladores. Un mismo neuromediador puede desempear cada
una de estas tres funciones.
Las neurohormonas son liberadas en una unin neurohemtica y actan lejos del sitio en
que fueron liberadas (GnRH, TRH, DA).
Los neurotransmisores clsicos actan en un espacio limitado de alrededor de 50 nm. Las
molculas neurotransmisoras liberadas en exceso son recapturadas en microsegundos por
protenas
especficas
(catecolaminas)
destruidas
por
degradacin
enzimtica
(acetilcolina).
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
Los neuromoduladores como indica su nombre, modulan la actividad o la sensibilidad de
las neuronas a un neurotransmisor [10].
Es muy frecuente que dos o ms neuromediadores se localicen en la misma neurona, lo que
constituye ms una regla que una excepcin. Como ejemplo se pueden citar, GnRH y
neurotensn; angiotensn y vasopresina; GABA y dopamina [10].
Hipfisis anterior y su neurorregulacin
La hipfisis anterior o adenohipfisis con frecuencia ha sido llamada la glndula maestra
por el lugar que ocupa en la jerarqua endocrina, ya que regula la actividad de distintos
rganos endocrinos, mediante las hormonas trpicas especficas que elabora para cada uno
de ellos.
Aunque a la hipfisis anterior le corresponde un lugar predominante entre las glndulas
endocrinas, descubrimientos de mediados del siglo indican que la actividad de esta a su vez
est regulada por el hipotlamo. Esta porcin del cerebro, como se dijo antes, produce y
secreta (bajo estmulos apropiados) factores de naturaleza polipeptdica que son
transferidos a travs de la circulacin portal hipofisaria a la hipfisis anterior, en este caso
una glndula subsidiaria, donde estimulan la secrecin de las hormonas hipofisarias.
Estas hormonas se forman en las clulas especficas de la adenohipfisis y permanecen
almacenadas en su protoplasma hasta que de algn modo los polipptidos hipotalmicos,
mediante una accin rpida, permeabilizan la membrana celular y hacen que salgan al
exterior [11].
Gonadotropinas hipofisarias
La LH y la FSH son las principales gonadotropinas de la pituitaria en la vaca. La prolactina
tiene un papel gonadotrpico en algunas especies, pero esto no ha sido aclarado en la vaca.
Antiguamente se crea que los centros hipotalmicos enviaban terminaciones secretoras al
lbulo anterior, y que estas terminaciones estimulaban la secrecin hormonal. Esta idea
pareca sustentada por el hecho de que la seccin del tallo hipofisario interrumpa, tanto el
celo como la secrecin lctea de los animales de experimentacin. Everett y Lawyer
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
descubrieron que esas rdenes secretoras partan efectivamente de los centros indicados,
pero no seguan una va nerviosa, sino una va hormonal.
La regulacin de la secrecin de gonadotropinas se cree que est en los tubrculos
mamilares y en el tuber cinereum, pero estos centros no emiten cilindro-ejes que terminen
en la adenohipfisis, sino que segregan sustancias que estimulan la secrecin en el lbulo
anterior. Estas sustancias, como se ha visto, alcanzan el lbulo por el sistema
portadienceflico que acarrea sangre desde la base del tercer ventrculo a la hipfisis, con
lo que la neurosecrecin de estos ncleos grises tienen ocasin de alcanzar el lbulo
anterior y excitar all la secrecin de sus clulas [11].
FSH. La hormona folculo estimulante (FSH) es una glicoprotena sintetizada por las
clulas basfilas de la hipfisis anterior y su vida media en la sangre de los bovinos es
aproximadamente de 5 h.
Un pico de FSH en el plasma coincide con la descarga preovulatoria de LH al inicio del
celo, con uno ms discreto aproximadamente 24 h ms tarde (Figura 3.7).
FIGURA 3.7 Niveles hormonales durante el ciclo estral de la vaca [1]
A lo largo del ciclo estral se producen picos adicionales al parecer relacionados con las
ondas de crecimiento folicular. En la vaca se ha descrito secrecin pulstil de esta
hormona. Recientes reportes sugieren que la FSH puede tener un papel lteo trpico [12].
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
La insulina aumenta el efecto estimulante de la FSH sobre las clulas de la granulosa, este
hecho, aunque an no se ha comprendido, tiene una gran significacin prctica, ya que las
vacas altas productoras (generalmente de baja fertilidad) tienen al principio del puerperio
muy bajos niveles de insulina [13].
L.H. La hormona luteinizante (LH) considerada como responsable de la maduracin y la
ovulacin del folculo de De Graaf y de la formacin y el mantenimiento del cuerpo lteo,
es tambin una glicoprotena, sintetizada por las clulas basfilas de la hipfisis a
semejanza de la FSH, pero a diferencia de aquella su actividad biolgica est representada
por la fraccin proteica y su vida media en sangre es solo de alrededor de 35 min.
Las concentraciones de LH son relativamente bajas durante la fase luteal del ciclo, pero
una descarga de esta en forma de un gran pico preovulatorio se produce de 24-30 h antes
de la ovulacin, esta coincide aproximadamente con el comienzo del celo.
En algunas especies hay evidencia clara de que la LH es liberada en episodios o pulsos a lo
largo del ciclo estral y en muchas est bien establecido que es necesaria la presencia de LH
para mantener la secrecin de progesterona por el cuerpo lteo.
Hipfisis posterior y su neurorregulacin
El lbulo posterior o neurohipfisis no es un rgano incretor, sus hormonas se originan en
los ncleos supraptico y paraventricular y emigran hasta dicho lbulo a lo largo de los
tractos nerviosos que conectan el hipotlamo con la glndula. Cuando se bloquean estos
tractos hay un acumulo de grnulos de secrecin en la parte hipotalmica de la zona
bloqueada, al tiempo que desaparecen gradualmente de la parte hipofisaria al irse vaciando
el lbulo neural de las hormonas almacenadas.
Estas hormonas son polipptidos, la ms importante para la reproduccin es la oxitocina, la
que produce contraccin de la musculatura lisa, principalmente del tero, pero tambin de
la vejiga, el intestino, el urter, la vescula biliar y la mama.
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
La oxitocina reduce de manera considerable la duracin del parto en la cerda y su empleo
est indicado particularmente en casos de inercia uterina, tanto en esta especie como en los
carnvoros.
La otra hormona es la vasopresina que tiene menos importancia directa sobre la
reproduccin. Su principal funcin es la regulacin del equilibrio acuoso en los animales,
tambin recibe el nombre de hormona antidiurtica [4]. Aunque ambas hormonas se
almacenan en el lbulo posterior de la hipfisis, los mecanismos que gobiernan su descarga
parecen ser diferentes.
La neurorregulacin de la secrecin de oxitocina se verifica a partir de tres centros
reflexgenos principales: el cuello uterino, la vagina y el pezn. Este reflejo se provoca de
un modo cruzado, tanto por la excitacin del pezn, como por la excitacin del cuello
uterino y probablemente tambin de la vagina [11]. Este reflejo muy rpido hace que una
excitacin de origen nervioso partida de estos puntos perifricos, vaya seguida de la
descarga de oxitocina que tiene tres misiones en este aspecto:
1. Contraer el tero en el momento del parto.
2. Ayudar a la eyeccin lctea y el vaciamiento de la ubre.
3. Ayudar al ascenso espermtico.
Durante el parto la dilatacin del cuello uterino y de la vagina provoca una descarga refleja
de oxitocina, que produce la mayor progresin del parto.
En el momento de la succin el ternero provoca la secrecin e igual sucede con el ordeo,
al estimular el pezn, por accin refleja de la hormona. Cuando se trata del recin nacido,
adems, aumentan las contracciones uterinas y los entuertos puerperales.
Se supone que el coito, la inseminacin artificial e inclusive las manipulaciones de la
vulva, dan origen a impulsos nerviosos que alcanzan el lbulo posterior de la hipfisis a
travs del hipotlamo y activan la descarga de la oxitocina, esta causa entonces las
contracciones uterinas y del oviducto, que son las responsables del rpido ascenso de los
espermatozoides desde el sitio en que fueron depositados hasta el oviducto. No existe
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
relacin entre los 21/2 min que demora el ascenso en la vaca y la velocidad de traslacin
de los espermatozoides con sus movimientos propios.
Funciones endocrinas del ovario
La actividad endocrina ovrica est representada principalmente por la produccin de tres
hormonas: dos esteroides (estrgenos y progesterona) y una proteica (relaxina); produce
otros esteroides, andrgenos y corticoesteroides, al parecer envueltos en el control del
desarrollo de los folculos y en el funcionamiento del cuerpo lteo. Recientemente se ha
descrito, adems, la produccin de oxitocina y de inhibina por el ovario.
Estrgenos
Los estrgenos como su nombre lo indica, son sustancias capaces de producir
manifestaciones de estro o celo en los animales. La hormona estrognica desarrolla los
caracteres sexuales secundarios en las hembras, por lo que es la hormona feminizante.
Las principales modificaciones morfolgicas producidas por los estrgenos en los rganos
genitales son: edema; hiperemia y crecimiento celular (epitelial y muscular). Los
estrgenos estimulan la contractilidad uterina y aumenta as la frecuencia y la amplitud de
sus contracciones. Accin semejante tienen sobre el oviducto.
El cuello uterino bajo la accin de los estrgenos segrega abundante moco y este se hace
ms fluido y ms fcilmente cristalizare (fenmeno de arborizacin) en el curso de su
desecacin.
Estas hormonas promueven la actividad fagocitaria del tero, estimulan el crecimiento del
epitelio vaginal y provocan su queratinizacin. La descamacin de clulas superficiales
aumenta. En el frotis vaginal los ndices cariopigntico y acidfilo se elevan.
Los estrgenos son galactogogos en hembras que no producen leche y galactoinhibidores
en las que la producen. En la vaca, en la cerda y en la perra son hipocalcmicos,
atribuyndoseles las frecuentes fracturas en vacas ninfmanas o sometidas a tratamientos
con estrgenos, y en las aves por el contrario tienen accin hipercalcmica.
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
En la coneja tienen accin abortiva hacia el final de la gestacin, y hasta un perodo
avanzado de esta en la vaca y la cabra; tambin ocurre en la perra, la gata y la rata, pero
solo antes de la implantacin.
En el macho los estrgenos se comportan como antagonistas de la testosterona.
El estrgeno de mayor actividad en la vaca es el 17 p estradiol, el que es sintetizado en los
folculos cavilarlos, en especial en los dominantes, por dos tipos de clulas. Las clulas de
la teca interna que sintetizan andrgenos a partir de colesterol y las de la granulosa que
sintetizan los estrgenos a partir de los citados andrgenos (Figura 3,8).
FIGURA 3.8 Representacin esquemtica de la sntesis de 17 p estradiol en los folculos ovricos [14]
La fuente principal de los estrgenos en la hembra no gestante es el folculo pero en la
gestante la cantidad mayor de esta hormona se produce en la placenta. Tambin es
producida en el testculo y en ambos sexos en cantidades muy pequeas en las adrenales.
Las concentraciones de estradiol son bajas a travs del ciclo estral, con tres picos que se
corresponden con las tres ondas de desarrollo folicular, el primero es un pequeo pico que
se produce alrededor del da 6 del ciclo, asociado con un folculo grande, cuando termina
la primera onda de desarrollo folicular; sobre el significado fisiolgico de este pico, se
especula acerca de su relacin con el transporte del huevo dentro del tero.
El segundo pequeo pico se produce a mediados del ciclo y se piensa que puede estar
relacionado con la induccin de receptores de oxitocina en el tero, para la subsiguiente
lutelisis.
Por ltimo, se produce un notable incremento de estradiol, que conforma un gran pico,
durante el perodo preovulatorio justamente previo o al comienzo del estro. Este es el
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
momento en que se ha incrementado el tamao del folculo dominante de la tercera onda
de desarrollo folicular, destinado a la ovulacin. El incremento de estradiol, estimula por
un mecanismo feedback positivo, la descarga preovulatoria de LH y FSH, que producen un
pico de 6- 8 h posteriores al de estradiol. Aproximadamente 1 h antes de la descarga de LH,
hay una declinacin notable de la concentracin de estradiol, la que aumenta de nuevo
acompaando a la descarga de LH (Figura 3.9).
FIGURA 3.9 Estradiol, progesterona y LH en una novilla tpica durante el ciclo estral [15].
Estas altas concentraciones de estradiol estn relacionadas con la mxima presentacin del
celo y con el comportamiento del toro. En las ovejas y posteriormente en las vacas, se ha
podido demostrar que la secrecin de estradiol est estrechamente acompaada de la
liberacin de un pulso de LH.
Los estrgenos como otras hormonas esferoides se segregan y pasan a la sangre como
esferoides libres, pero rpidamente una parte se convierte en formas ligadas por
conjugacin con glucuronato o con sulfato, o se une a las protenas. Los estrgenos
circulantes unidos a las protenas tambin parecen estar conjugados con el cido
glucurnico. Estos procesos tienen lugar en el hgado.
Se crea que la formacin de conjugados constitua una etapa del proceso de excrecin,
pero en la actualidad se piensa que puede ser un paso importante en la funcin de la
hormona. Los estrgenos son fisiolgicamente activos en concentraciones mucho menores
que los otros esferoides y son capaces de antagonizar los efectos biolgicos de grandes
cantidades de otras hormonas.
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
El estradiol que circula por la sangre es inactivado en el hgado, con formacin de estrena,
estril y cidos biliares. La estrena y el estril son ms fcilmente eliminables por el rin
que el estradiol, por lo que el hgado no solamente contribuye a inactivar en parte los
estrgenos circulantes, sino que favorece de manera indirecta su eliminacin [11]. Los
estrgenos se eliminan en cantidades apreciables por la bilis. Ya en el intestino el 40 % se
recupera por el emuntorio intestinal y el resto es eliminado por las heces. El paso a travs
de la va entero heptica reduce considerablemente el poder de los estrgenos naturales,
por lo que su administracin por va digestiva debe ser descartada.
Existen varios compuestos sintticos con actividad estrognica, pero el ms utilizado es el
dietil estilbestrol o estilbestrol,
Progesterona
La progesterona siempre es producida por el cuerpo lteo al principio de la preez y es
esencial para el mantenimiento de la gestacin. Tambin se produce en la placenta de
algunos animales, pero en determinadas especies (yegua y oveja) al final de la gestacin
dicha produccin alcanza gran importancia, al sustituir a la producida por el cuerpo lteo.
Tambin se encuentra progesterona, en pequeas cantidades, en la corteza adrenal y en el
testculo, probablemente por ser intermediaria de los corticoides adrenales y de la
testosterona.
En los carnvoros la placenta no secreta progesterona, excepto en la gata al final de la
gestacin [16].
La progesterona hormona de la preez es un factor de primera necesidad para el
mantenimiento de la gestacin. Despus de producida la fecundacin, esta hormona inhibe
la actividad contrctil del tero y estimula el desarrollo de sus glndulas. Tambin provoca
la mucificacin del epitelio vaginal y ejerce accin hiperplsica sobre los acinis
glandulares de la mama.
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
Tanto las clulas luteales grandes como las pequeas del cuerpo lteo secretan
progesterona, pero las pequeas responden a la LH in vitro, produciendo seis veces ms
progesterona que las grandes [17].
Esta hormona ejerce un efecto feedback negativo sobre la liberacin de LH, aparentemente
por reducir la frecuencia de los pulsos de LH. La progesterona tambin es secretada en
forma pulstil, los pulsos coinciden durante la fase luteal con los de FSH [12] ms bien que
con los de LH. De este modo la evidencia de que la LH es la mayor luteotropina puede
requerir un reexamen.
El cuerpo lteo de la oveja alcanza el mximo de secrecin de progesterona alrededor del
da 7 del ciclo, y sobre el 15, un da antes de manifestarse el celo, esta declina
abruptamente (ver Figura 3.9). Esto es el disparador de la secuencia de cambios
hormonales que produce el nuevo estro y la ovulacin. Si el cuerpo lteo regresa
prematuramente el animal vuelve al celo antes de lo esperado; si la vida del cuerpo lteo se
prolonga el estro es pospuesto. Por tanto, el cuerpo lteo es el reloj que controla la
duracin del ciclo estral [18].
El mecanismo de secrecin de la progesterona no es conocido. No ha sido aclarado si
ocurre por difusin pasiva del esteroide a travs de la membrana plasmtica o por
transferencia despus de unirse una molcula de protena o de lpido [19]. Gran parte de la
progesterona circulante es probable que se encuentre conjugada en forma de cido
glucurnido.
Al igual que otros esteroides, esta hormona no se almacena en el organismo, siendo
excretada despus de sufrir un proceso de reduccin, en el que desempea un papel
importante el hgado. En casi todas las especies animales el principal producto de
excrecin de la progesterona es el pregnandiol, el que aparece en la orina en estado de
glucurnido. Como esta hormona desaparece rpidamente de la circulacin general, es
necesaria que su destruccin sea compensada por una produccin constante. La cantidad de
hormona secretada por una vaca durante la gestacin puede ser de 100-300 mg diarios.
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
Relaxina
La relaxina considerada la segunda hormona del cuerpo lteo, es un polipptido
hidrosoluble, descubierto por Hisaw en 1926, capaz de provocar la sinfilisis en la cobaya
y la rata. Al principio su naturaleza hormonal fue negada por algunos. El descubrimiento
de la progesterona oscureci por su importancia al de la relaxina, retrasando su estudio. El
haber llegado al conocimiento de esta sustancia olvidada es el resultado de la tenacidad y
la conviccin cientfica de Hisaw y sus colaboradores.
Se puede extraer relaxina de los ovarios de vacas gestantes [20] y se detecta en la sangre al
final del primer mes de la gestacin. Los niveles entonces aumentan hacia el final del
segundo tercio y despus permanecen constantes hasta despus de las 24 h posteriores al
parto, cuando ocurre una rpida disminucin.
Se ha comprobado que esta hormona causa despolimerizacin del tejido conectivo y tiene
un significativo papel en la dilatacin del crvix en la vaca y en la cerda, as como en la
relajacin de los ligamentos sacrociticos al momento del parto. Este efecto se potencializa
con el pretratamiento con estrgenos y progesterona. En el futuro la relaxina se podr usar
para reducir la incidencia de dificultades en el parto, especialmente en vacas pequeas
[13].
La relaxina es sintetizada y secretada por el cuerpo lteo durante la gestacin en rumiantes,
cerda, rata y mujer, por el endometrio en la coneja y la yegua, y por la placenta en la
cobaya y la mujer [21].
Aunque dicha hormona es considerada una hormona de la gestacin, se encuentra tambin
presente en grandes cantidades en la sangre de cerdas jvenes y adultas no preadas,
considerndose a las clulas de la granulosa como su posible fuente. Su funcin en
animales no preados se desconoce [1].
Oxitocina
Se ha encontrado oxitocina en el cuerpo lteo de las ovejas. La inmunizacin activa contra
la oxitocina prolonga la vida del cuerpo lteo de esta especie, lo que ha sugerido que la
oxitocina puede estar envuelta en el proceso de lutelisis. Abunda en este sentido el hecho
de que su contenido disminuye en el cuerpo lteo despus de la fecundacin [22].
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
El papel de la oxitocina en el ciclo estral de los bovinos se desconoce, pero se han
demostrado pulsos de esta en la vena cava posterior que coincide con los de progesterona
[12].
Hoy da se cree que la oxitocina luteal viaja al tero, donde causa liberacin de
prostaglandina F2 a. Esta estimula nueva liberacin de oxitocina luteal en forma de
cascada de hormonas por el ovario y el tero en el apropiado momento del ciclo [13]. Si la
fase luteal se prolonga, por ejemplo en los casos de gestacin, el cuerpo lteo no contiene
ms oxitocina y pierde la habilidad de sintetizarla [21].
Inhibinas
Las inhibinas son una familia de hormonas pptidas capaces de regular parcialmente la
secrecin de FSH, estimulando o inhibiendo su respuesta al gonadotropin releasing hormn
(GnRH). Estas han sido aisladas de extractos ovricos y testiculares [23].
En la hembra, las inhibinas son sintetizadas por las clulas de la granulosa. La hormona
sintetizada se acumula en el lquido folicular, encontrndose grandes concentraciones en el
fluido de los folculos grandes y medianos, o es secretada dentro del plasma [23]. Las
inhibinas son secretadas en discretos pulsos sincronizados con los pulsos de progesterona y
de LH [9].
Actan a nivel de la pituitaria donde se-han demostrado receptores para las mismas. La
adicin de inhibinas a cultivos de clulas de la pituitaria est acompaada por un 60-75 %
de disminucin en la secrecin de FSH. La dinmica de esta reaccin es lenta. Bajo
condiciones experimentales similares, la secrecin basal de LH no es modificada [9].
Tambin estas hormonas reducen la secrecin de FSH inducida por la GnRH. Usualmente
no tienen o tienen un efecto menor sobre la secrecin de LH inducida por la GnRH. Las
ovejas inmunizadas contra las inhibinas obtenidas de folculos bovinos, tienen un
incrementado nmero de ovulaciones [24].
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
Las inhibinas estn constituidas por dos cadenas de naturaleza peptdica unidas por lazos
de sulfuro. Las cadenas han sido denominadas: alfa, beta A y beta B. Las que reducen la
liberacin de FSH son, la inhibina A y la inhibina B. La primera est constituida por una
cadena alfa unida a una beta A y la segunda por una cadena alfa pero unida a una beta B.
Cuando las dos subunidades estn unidas el efecto es el opuesto al de inhibicin. Estas
adivinas que estimulan la secrecin de FSH son las adivina A y la adivina A-B. La adivina
A formada por dos cadenas beta A y la adivina A-B por dos cadenas beta, las beta A y B
(Tabla 3.1). La adivina es el verdadero FSH-RH, que se busc durante muchos aos
intilmente en el hipotlamo, y que al fin en 1985, encontraron Vale y Guillemn,
simultnea e independientemente en ambas gnadas [25]. Esta molcula, llamada adivina o
FRP (FSH releasing protein) estimula la secrecin de FSH por las clulas de la hipfisis
pero no influye a la LH [26].
TABLA 3.1 COMPOSICIN Y ACTIVIDAD DE LAS INHIBINAS
Inhibina
Cadenas
Actividad
Inhibina A
alfa-beta-A
Inhibe secrecin FSH
Inhibina B
alfa-beta B
Inhibe secrecin FSH
Activina A
beta A-beta A
Estimula secrecin FSH
Activina A-B
beta A-beta B
Estimula secrecin FSH
Recientemente se ha demostrado en la oveja preada, un factor de inhibicin de liberacin
de la hormona luteinizante (LH-RIF). Es detectable desde el da 15 de la gestacin y su
nivel es elevado entre los das 70 y 120, pero est ausente en el cuerpo lteo cclico. En
contraste con las inhibinas y adivinas, este LH-RIF no afecta la secrecin de FSH pero
inhibe los pulsos de LH.
Andrgenos
Se encontraron apreciables cantidades de testosterona (2,6 ng/g de tejido) en el cuerpo
lteo de la vaca, el da 12 del ciclo estral [27]. Posteriormente Wise et al. [28] mostraron
que las concentraciones de androstenodiona, dehidroepiandrostenediona y testosterona
aumentan en la vena ovrica al aproximarse el estro, lo que seala su origen ovrico.
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
El papel de los andrgenos en la hembra no est claro, pero se sabe que estos son
intermediarios en la biosntesis de otros esferoides como los estrgenos. No obstante, es
posible que puedan desempear algn papel en el control de la secrecin de
gonadotropinas y en la induccin del comportamiento estral [29].
Tambin existe la posibilidad de que la PGF2a coordine el crecimiento folicular y la atresia
con la muerte del cuerpo lteo, a travs de andrgenos luteales, ya que se han observado
incrementos en testosterona ovrica dentro de un perodo de 4 das antes de la regresin del
cuerpo lteo [28].
Mecanismo feedback
Se postularon tres mecanismos feedbacks hormonales [7], actuando sobre el eje
gonadotrpico, uno largo, desde el ovario al hipotlamo, uno corto desde la hipfisis al
hipotlamo y uno ultracorto en el propio hipotlamo (Figura 3.10).
FIGURA 3.10 Tres mecanismos de retroaccin hormonal (feedbacks) [30].
Feedback de los esteroides gonadales sobre el hipotlamo {feedback largo)
Los esteroides gonadales son los principales reguladores de la secrecin de GnRH [14]. La
primera accin del estradiol, la progesterona o la testosterona, es disminuir la frecuencia de
generacin de los pulsos de GnRH, no la de modificar su biosntesis [31]. Despus de un
primer perodo de inhibicin (retroaccin negativa) el estradiol incrementa durante la fase
folicular la secrecin de GnRH (retroaccin positiva), la que induce una descarga
preovulatoria de LH [5].
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
Feedback hipofisario (feedback corto)
La hiptesis de que exista un feedback corto desde la hipfisis al hipotlamo a travs del
cual la secrecin de LH est involucrada en el control de la secrecin de GnRH, se basa en
la existencia de una corriente sangunea retrgrada en los vasos del sistema portal
hipofisario. Pero un bloqueo total de la secrecin de LH, o un gran incremento de los
niveles de LH mediante la administracin de esta hormona, no tiene efecto en los
parmetros de secrecin de GnRH, lo que no sostiene la existencia de un feedback corto,
por el que la LH inhiba la secrecin de GnRH [5].
Feedback hipotalmico (feedback ultracorto)
La administracin (perifrica o intraventricular) de grandes cantidades de GnRH o de un
antagnico de dicho pptido, puede conducir a una reduccin de la secrecin de GnRH. No
obstante, el papel de un feedback hipotalmico como un posible regulador de los pulsos de
GnRH, todava no ha sido demostrado en condiciones fisiolgicas [5].
Factores fisiolgicos que controlan al cuerpo lteo
Dada la importancia del cuerpo lteo en el ciclo ostral, es necesario examinar los factores
fisiolgicos que controlan esta transitoria glndula endocrina.
Cuando se produce la ovulacin, los vasos sanguneos de la membrana basal del folculo se
rompen. La cavidad del folculo roto es infiltrada por eosinfilos, las clulas de la
granulosa y de la teca interna se hipertrofian, proliferan y comienzan a luteinizarse, as
como su citoplasma crece y un suave retculo endoplsmico empieza a desarrollarse para
constituir las clulas luteales que van a formar el cuerpo lteo. Esta glndula de 2 - 4 das
despus de producirse la ovulacin, sintetiza y secreta apreciables cantidades de
progesterona. Las clulas de la granulosa son conocidas como grandes clulas luteales y las
de la teca como pequeas clulas luteales.
Existen marcadas diferencias en las respuestas de estas clulas a los estmulos hormonales.
Las pequeas clulas luteales incrementan su produccin de progesterona en respuesta a la
LH y no responden en ese sentido a la prostaglandina E, mientras que las grandes no
responden al estmulo de la LH, pero s al de la prostaglandina E. En contraste con esto,
dichas clulas luteales responden a la prostaglandina F, disminuyendo su produccin de
progesterona, y las pequeas le dan una respuesta menor a la prostaglandina.
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
Hay evidencia de que algunas pequeas clulas luteales pueden transformarse en grandes
clulas cuando el ciclo estral progresa, proceso que aparentemente contina durante la
gestacin [13]. Tambin existe evidencia de que las grandes clulas luteales pueden
producir una sustancia citotxica que destruye a las pequeas durante la lutelisis.
En los primates las clulas luteales son inmediatamente funcionales y secretan
incrementadas cantidades de progesterona desde el da de la ovulacin. En los rumiantes el
cuerpo lteo comienza a funcionar despus de uno o dos das, esta demora puede estar
relacionada con los; grandes cambios histolgicos que ocurren en estas especies.
La vida del cuerpo lteo de ciclo es variable de especie a especie, usual-mente dura entre
12-21 das, pero es constante ( un da) para cada especie. La variacin en concentraciones
de progesterona en sangre a travs de la fase luteal refleja los sucesivos estadios de
crecimiento, mantenimiento y regresin del cuerpo lteo [19].
La transicin de un folculo secretor de estrgenos en un cuerpo lteo secretor de
progesterona est dada, porque (as clulas de la teca interna del, folculo antes de
producirse la ovulacin poseen un sistema enzimtico, que transforma la progesterona en
andrgenos y estos en estrgenos, pero cuando dichas clulas se luteinizan como
consecuencia de la ruptura del folculo, son relativamente deficientes en ese sistema
enzimtico [18].
Actividad luteoltica del tero
A partir de la observacin de Leo Loeb, de que la histerectoma en la cobaya prolongaba la
vida del cuerpo lteo, distintos grupos de investigadores comprobaron que el tero ejerce
una accin ltica sobre este. Demostraron que si se extraa el tero de una oveja, que
tuviera un cuerpo lteo floreciente, la vida de esta glndula que normalmente es de 15 das,
se prolongaba por varios meses (Figuras 3.11 y 3.12).
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
FIGURA 3.11 Oveja normal, el cuerpo lteo dura 15 das [32].
FIGURA 3.12 Oveja histerectomizada, el cuerpo lteo dura vahos meses [32].
Comprobaron tambin que la influencia ltica operaba en forma local, ya que cuando se
extraa el cuerno adyacente al ovario que tena un cuerpo lteo, este prolongaba la vida por
muchos das, pero si se extraa el prximo al ovario que no tena cuerpo lteo, el mismo
tena una duracin normal de ', o das (Figuras 3.13 y 3.14).
FIGURA 3.13. Al extraer el cuerno prximo al cuerpo lteo su funcin se prolonga muchos das [32]
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
FIGURA 3.14. Al extraer el cuerno prximo al ovario sin cuerpo lteo la vida de la glndula es de 15 das
[32]
En los animales que tienen tero unicrneo, por aplasia del otro cuerno, el comportamiento
es similar, dependiendo la vida del cuerpo lteo del ovario en que se produjo la ovulacin.
As, si el cuerpo lteo se encuentra en el lado que no est el cuerno, falta la accin ltica, y
dura muchos das, mientras que si se encuentra en el lado que existe el cuerno se produce
dicha accin y el cuerpo lteo tiene su duracin normal de 15 das (Figuras 3.15 y 3.16).
FIGURA 3.15 tero unicrneo, si la ovulacin se produce en el ovario correspondiente al cuerno aplsico, la
vida de] cuerpo tuteo se prolonga por muchos das [32].
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
FIGURA 3.16 tero unicrneo, si la ovulacin se produce en el ovario correspondiente al cuerno presente, la
vida del cuerpo lteo es de 15 das [32].
As parece que hay un ciclo tero-ovrico y que posiblemente el cuerpo lteo estimule al
tero a producir una sustancia que con posterioridad lo destruir. Por muchos aos se ha
conocido que el tero produce una hormona luteoltica que causa la regresin del cuerpo
lteo al final del ciclo estral. Ya en 1959, Armstrong y Hansel [33] mostraron la accin
luteoltica o al menos antiluteotrpica de la oxitocina exgena, en la vaca.
Ginther et al., en 1967 [34] observaron que ese efecto no se originaba si el animal era
histerectomizado, o si se le extraa el cuerno ipsilateral al ovario que tena el cuerpo lteo.
En 1972, Harrison et al. [31] y Me Cracken et al. [35] mediante la determinacin de
prostaglandina F2a, en el fluido uterino de ovejas, a las que le haban trasplantado el ovario
demostraron que era esa la sustancia luteoltica. La prostaglandina F2a y sus anlogos
sintticos son potentes agentes luteolticos en la vaca y se emplean en ellas ampliamente
con ese propsito.
Hoy da se cree que la prostaglandina F2a del tero es secretada en forma pulstil y que
estos pulsos son dirigidos por pulsos de oxitocina desde el cuerpo lteo. La habilidad de la
oxitocina para estimular la secrecin de prostaglandina F depende de los receptores de
oxitocina del tero. A mediados del ciclo los estrgenos producidos por el folculo
dominante de la segunda onda de crecimiento folicular inducen la formacin de receptores
de oxtocina en el tero. Su produccin se regula por los niveles de progesterona; cuando
esta aumenta, disminuye el nmero de receptores, pero si comienza a declinar, el nmero
de receptores aumenta. Cuando estos son ocupados por la oxitocina, activan la fosfolipasa
A2 y liberan cido araquidnico, la cascada de este cido es la principal productora de
PGF2a uterina y eventual lutelisis [36].
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
La prostaglandina F2a aparentemente estimula ms liberacin de oxitocina del cuerpo
lteo y esta cascada contina hasta que la lutelisis es completa (Figura 3.17). Muchos
factores, incluyendo la infeccin uterina, al parecer interfieren con este proceso y provocan
la prolongacin del ciclo estral o ciclos irregulares [13].
FIGURA 3.17 Estimulacin recproca de secrecin de oxitocina (por el cuerpo lteo) y de PGF 2 (por el
tero) en la oveja: A. La inyeccin intravenosa de PGF 2 causa un aumento inmediato del nivel de oxitocina
en la sangre de la vena tero ovrica (VUO) y en la vena perifrica (VP). B. La inyeccin intravenosa de
oxitocina causa un aumento inmediato de PGFM (metabolito estable de la PGF 2) en una vena perifrica
[37].
La lutelisis no puede ser en ninguna forma atribuida a cambios en el plasma de los niveles
de hormonas luteotrpicas, como la LH y la prolactina. Adems, en todos los mamferos,
una disminucin en el nmero de pulsos de GnRH ocurre durante la fase luteal;
inyecciones pulstiles de GnRH o de LH con un ritmo constante no pueden interrumpir la
lutelisis [38].
PROSTAGLANDINAS
Las prostaglandinas son cidos grasos no saturados derivados del cido prostanoico. Se
encuentran en todo el organismo animal. Se han identificado alrededor de 15 series de este
tipo de hormona con las ms diversas funciones, y se conoce el papel que desempean en
la reproduccin las series F y E. Son consideradas como hormonas locales debidos a que
tienen tendencia a ser inestables y rpidamente metabolizadas.
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
La vida media en sangre de la prostaglandina F2a es aproximadamente de 8 min. y en los
pulmones es metabolizada inicialmente a 13,14-dihydro-15-keto-PGF2a (PGFM). Este
metabolito ms estable es usado para monitorear cambios en los patrones de liberacin ms
estable de esta hormona.
Pulsos luteolticos de PGF2a (PGFM perifrica) son observados con concurrentes
incrementos de estradiol y declinantes concentraciones de progesterona [39]. As se ha
encontrado que las concentraciones de PGFM en el plasma perifrico son bajas la mayor
parte del ciclo estral y que sufren un incremento en el momento de la regresin luteal [40].
La forma en que la PGF2a, de corta vida, producida en el tero, puede llegar al cuerpo
lteo, para ejercer su funcin luteoltica era desconocida hasta que Goding en Australia
separ meticulosamente la arteria y la vena ovrica que estn estrechamente adheridas. Las
ovejas, a las que se les separ fallaron en presentar lutelisis.
Esto le permiti formular la teora de que existe un mecanismo contracorriente que permite
que una sustancia luteoltica del tero pase de manera directa desde la vena tero-ovrica a
la arteria uterina y de este modo pueda llegar al cuerpo lteo [41] (Figura 3.18).
FIGURA 3.18 Transferencia de la prostaglandina F2a del tero al ovario mediante un sistema contracorriente
[42].
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
Mecanismo de la lutelisis
No est claro el mecanismo por el cual la PGF2a induce lutelisis. Su efecto luteoltico se
piensa ocurre, en parte a travs de una intervencin directa sobre las clulas luteales, e
indirecta por medio de la reduccin de la irrigacin sangunea al cuerpo lteo, como
sugirieron Pharris et al, en 1970 [43S Aunque la PGF2a tiene propiedades
vasoconstrictoras, no se ha determinada si la marcada disminucin de la corriente
sangunea en el ovario o en el cuerpo lteo, o en ambos, la que ocurre en lutelisis
espontnea o inducida con PGF 2a, es la causa o la consecuencia de la lutelisis [39].
La forma en que la prostaglandina F 2a ejerce su accin luteoltica ha sido objeto de
controversias. Pharris et al., en 1970 [43], sugirieron que la lutelisis es causada por una
restriccin del flujo sanguneo al cuerpo lteo, pero es posible que esto sea consecuencia
ms que causa de la regresin. Posteriormente se ha sugerido que esta hormona interfiere
con la unin de la LH y la adenilciclasa de la membrana celular.
Mecanismo de regulacin neurohormonal del ciclo estral
Las relaciones hipotlamo-hipfisis-ovario-tero, de acuerdo con los conocimientos
actuales se pueden esquematizar del modo siguiente: bajo la accin del factor de liberacin
gonadotrpico hipotalmico, la hipfisis libera las hormonas FSH y LH necesarias para
provocar las ondas de crecimiento folicular, que concluyen con la maduracin de uno o
varios folculos (segn la especie); esta maduracin folicular se acompaa de la secrecin
de estrgenos que estimula por un mecanismo feedback positivo la descarga preovulatoria
de LH y FSH.
Los estrgenos producidos en la segunda onda de crecimiento folicular inducen la
formacin de receptores de oxitocina en el tero. De 2-4 das despus de producirse la
ovulacin el cuerpo lteo formado en la cavidad del folculo roto comienza a secretar
cantidades apreciables de progesterona en respuesta a estmulos recibidos de LH en las
pequeas clulas luteales y de prostaglandina E en las grandes clulas luteales.
Los niveles elevados de progesterona ejercen un efecto feedback negativo sobre la
liberacin de LH posiblemente reduciendo la frecuencia de sus pulsos. Mientras tanto, en
el cuerpo lteo, se origina tambin oxitocina, probablemente en las grandes clulas
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
luteales, la que actuando sobre los receptores del tero estimula la liberacin de
prostaglandina F2a, de accin luteoltica. La PGF2a al parecer estimula ms secrecin de
oxitocina del cuerpo lteo, formando una cascada que provoca un cambio brusco de los
niveles de progesterona, hasta alcanzar niveles bsicos, lo que va a actuar como disparador
del prximo ciclo.
Gonadotropinas extrahipofisarias
En la orina de las mujeres y en la sangre de las yeguas y burras gestantes se hallan
sustancias gonadotrpicas de accin bastante semejante a la de las gonadotropinas
hipofisarias. El factor gonadotrpico presente en la orina de mujeres gestantes tiene una
fuerte accin luteinizante, sin embargo, en la gonadotropina srica que se encuentra en las
equinas preadas, la accin fundamental es folculo estimulante. Estas caractersticas de las
gonadotropinas extrahipofisarias, las hace adquirir gran importancia, al constituir una
fuente prctica y econmica de su empleo en medicina humana y veterinaria debido a su
disponibilidad y precio menos elevado.
Gonadotropina srica PMSG o gonadotropina corinica equina eCG
Harold Col, en 1930 descubri en la sangre de yeguas preadas altos niveles de
gonadotropina, la que tiene accin estimulante sobre los folculos ovricos y las clulas
intersticiales del testculo y poca actividad luteinizante. Esta gonadotropina es una
glicoprotena de gran peso molecular aproximadamente 70 000 Dalton, por lo que no
aparece en la orina como la gonadotropina extrahipofisaria de la mujer.
La gonadotropina srica (PMSG) pregnantmare serum gonadotrophin, en la actualidad
llamada gonadotropina corinica equina (eCG), por originarse en el trofoblasto que cubre
los clices endometriaies, aparece en la sangre de las hembras equinas (yegua y burra)
entre 35 y 40 das del comienzo de la gestacin, y alcanzan grandes concentraciones entre
los 50 y 90 das, con la mxima concentracin entre 60 y 70, manteniendo pequeas
cantidades hasta los 140o 150 das.
Las razas ligeras y las ms pequeas producen, en general ms PMSG por litro de suero
que las razas pesadas, por lo que se encuentran diferencias individuales notables.
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
La gonadotropina srica se produce en los clices endometriales (endometrial cups), por lo
que se consideraba de procedencia maternal, pero Allen y colaboradores demostraron que
esas estructuras son de origen fetal, por lo que hoy se reconoce la naturaleza
fetoplacentaria de esta gonadotropina [32].
Alrededor del da 35 de la gestacin se comienzan a desprender clulas de una banda
ecuatorial del alantocorion que se ha desarrollado y rodea al embrin. Estas clulas
penetran en el endometrio materno horadando profundamente su estroma en el que se
agrandan para formar las caractersticas clulas decidualesde los clices endometriales.
Cerca del da 40 el tejido de los clices se hace visible a simple vista y aparece en la
circulacin materna la PMSG.
El nivel de gonadotropina aumenta y alcanza el mximo alrededor del da 60.
Aparentemente la madre ante la invasin de su estroma endometrial desarrolla una clsica
respuesta inmunolgica para rechazarla. Las clulas deciduales son rodeadas por una masa
de linfocitos y posiblemente el tejido de los clices sea desprendido de la superficie del
tero al interior del lumen. Cuando los linfocitos rodean a las clulas deciduales la PMSG
comienza a declinar y cae bruscamente si el tejido de los clices se desprende del tero.
En general, se dice que los tejidos fetales son protegidos completamente del ataque
inmunolgico maternal, por los que los clices endometriales de la yegua proveen una
intrigante excepcin a esa regla.
En contraste con los primates la eCG no contribuye al mantenimiento del cuerpo lteo al
principio de la gestacin, porque la eCG comienza a secretarse mucho tiempo despus de
la inhibicin de la regresin del cuerpo lteo. La eCG est envuelta en la
inmunorregulacin de la implantacin que tiene lugar desde el da 35 en la yegua [16]. Esta
hormona fue muy usada para el diagnstico de la gestacin, pero, la forma esfrica del
conceptus, en esta especie, permite ahora establecer un rpido diagnstico por ecografa
(desde el da 10 de la gestacin).
En la yegua, al principio de la gestacin se producen incrementos de la secrecin de FSH a
intervalos de aproximadamente 10 das, esto resulta en ondas de actividad folicular. La
secrecin de cantidades masivas de gonadotropina corinica equina que comienza
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
alrededor del da 35 de la gestacin induce la ovulacin de estos folculos y la formacin
de cuerpos lteos secundarios.
La funcin del cuerpo lteo primario tambin se estimula mediante la gonadotropina
corinica equina. Los cuerpos lteos secundarios son similares al cuerpo lteo primario en
estructura y funcin. Estos secretan progesterona hasta que su soporte (eCG) desaparece de
la circulacin ms o menos el da 150 de la gestacin en que ellos regresan junto con el
cuerpo lteo primario. El soporte progestacional para el resto de la gestacin es provisto
por la unidad fetoplacentaria [44]. Una gonadotropina corinica ha sido identificada, pero
no purificada en rumiantes (vaca y oveja), en la rata, la ratona y la coneja [16].
Gonadotropina corinica humana (hCG)
La gonadotropina corinica, human chorionic gonadotrophin (hCG), es tambin una
glicoprotena de peso molecular alrededor de 30 000 Dalton, se halla en la orina y el suero
sanguneo de las mujeres gestantes y tambin en la orina de las hembras del Chimpanc y
del Macaco rhesus. Su estructura qumica y propiedades son muy similares a las de la
hormona LH, aunque tiene un mayor contenido de carbohidratos, por lo que es
relativamente resistente a la degradacin metablica, de aqu que tenga una vida media
biolgica mucho mayor que la de la LH [45].
Su presencia en la orina de mujeres gestantes fue utilizada por Zondek y Ascheim, en
1927, para el diagnstico precoz de la gestacin, al comprobar que provoca la ovulacin y
hemorragias en los folculos ovricos, cuando se inyecta a ratones, conejos, ratas y sapos.
La gonadotropina corinica es producida en el sincitio y el citotrofoblasto de las
vellosidades coriales. Tambin se originan en grandes cantidades en las molas
hidatiformes, en el corioepitelioma maligno y en un raro tumor testicular que contiene
tejido trofoblstico. Su presencia en la orina se puede detectar ya desde el da 16 posterior
a la fecundacin, y alcanza un pico aproximadamente entre los 30 y 60 das, despus
desciende y mantiene bajos niveles durante el resto de la gestacin.
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
Propiedades antignicas de las gonadotropinas
Las gonadotropinas tienen propiedades antignicas, lo que provoca un estado refractario a
ellas en los animales que son sometidos a tratamientos repetidos con gonadotropinas
extraas o heterlogas.
Feromonas sexuales
Las feromonas sexuales son sustancias odorferas producidas comnmente en las glndulas
modificadas de la piel y constituyen el medio de identificacin de la disposicin sexual de
los animales y les permite reconocer a considerables distancias a las hembras en celo, as
como ubicarlas dentro del rebao. Las hembras, a su vez, son estimuladas sexualmente por
las feromonas del macho de su especie.
Aunque las feromonas no se ajustan a la definicin de hormonas de Starling, deben
reconocerse como miembros en el concepto ms amplio de una hormona o mensajero
qumico externo [13]. Determinadas feromonas pueden ser reconocidas por el olfato
humano, como las producidas por el macho cabrio y por el verraco.
Reconocimiento de la gestacin por la madre
Para que se desarrolle la gestacin es indispensable mantener el cuerpo lteo funcionando
y suprimir la ciclicidad estral. Como el organismo animal reconoce que se ha producido la
fecundacin del huevo, que ha comenzado una gestacin y que, por lo tanto, debe
prolongarse mientras dure esta, la fase luteal del ciclo estral, es una cuestin de sobrada
importancia y no completamente aclarada.
La yegua, especie en la que solo los huevos fertilizados pueden pasar al tero, muestra que
debe existir un mecanismo extremadamente rpido, para reconocer la preez en estos
animales. En otras especies la blstula debe dar la seal de que se ha producido la
gestacin, despus de haber penetrado en el tero, pero antes de su implantacin.
En la oveja si el da 12 del ciclo se le transfiere un embrin de 12 das en un cuerno
uterino, el cuerpo lteo de ciclo se transforma en un cuerpo lteo de gestacin. El efecto
antiluteoltico inducido de este modo por el conceptus es el resultado de una accin local
sobre el tero (Figura 3.19). Verdaderamente, cuando el cuerno uterino en que el
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
blastocisto es transferido, es aislado por una ligadura, la lutelisis solo ser inhibida en el
caso en que el cuerpo lteo est situado en el lado del cuerno grvido [46].
FIGURA 3.19 Papel antiluteoltico local del embrin en la oveja. Embrin de 12 das transferido al cuerno
uterino el da 12 del ciclo estral [19].
Si no hay blastocisto en el tero de la oveja el da 12, en la cerda el 13 y en la yegua entre
los das 14 y 16 del ciclo, el cuerpo lteo empieza a degenerar [47]. En la cerda se requiere
un mnimo de cuatro embriones para mantener la gestacin [48].
Cuerpo lteo de gestacin
La transformacin de un cuerpo lteo de ciclo en un cuerpo lteo de gestacin requiere de
la inhibicin de la lutelisis y del mantenimiento del estimulo hormonal luteotrpico.
En los primates las dos acciones son realizadas por la gonadotropina corinica, pero en
muchas otras especies, incluyendo los rumiantes, estas acciones estn separadas. En estos
la transformacin de un cuerpo lteo de ciclo en uno de gestacin es lograda por la
intervencin del conceptus, que inhibe la accin luteoltica del tero, al disminuir en
magnitud y frecuencia la secrecin pulstil de PGF2a por el endometrio (Figura 3.20) y por
el mantenimiento de la accin luteotrpica de la hormona hipofisaria LH con una
contribucin progresiva de importancia variable, de la placenta [19].
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
FIGURA 3.20 Progesterona en la oveja y PGF 2. Evolucin paralela de niveles del principal metabolito de
la PGF 2 (PGFM = 13,14 dihydro 15 keto PGF2a) y de la progesterona durante el ciclo estral y el
principio de la gestacin en la oveja [49]
Recientemente se ha determinado que en el ganado alrededor del da 15o 16 despus del
celo, el conceptas produce la liberacin de una protena, la trofoblstica bovina 1 (btp-1)
que acta sobre el tero reduciendo la cantidad de prostaglandina F2a que este rgano
libera. La btp-1 no se detecta en sangre, orina ni moco cervical, estando su localizacin
limitada al cuerno uterino gestante. Esta protena puede bloquear los receptores de la
oxitocina del tero y de este modo bloquear la habilidad de la oxitocina de liberar
prostaglandina F2a [13].
El efecto antiluteoltico se prolonga luego de que ha sido disparado por la presencia de un
embrin en el tero. Despus de eliminado el conceptas por un lavado uterino entre los
das 21-23 de la preez, el cuerpo lteo puede permanecer funcional por muchos meses
[50]. Esta persistencia del cuerpo lteo debe ser explicada por la desaparicin de la sntesis
de oxitocina en el cuerpo lteo de gestacin y la baja concentracin de receptores de
oxitocina.
La inyeccin intrauterina de trofoblastina natural o recombinante en ovejas cclicas puede
mantener el cuerpo lteo funcional por ms de un mes (Figura 3.21). La trofoblastina
puede, adems, ser considerada como un interfern del embrin expresado, naturalmente,
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
en ausencia de infeccin viral. La protena trofoblstica ovina (oTP) purificada exhibe la
actividad antiviral caracterstica de los interferones (IFNs) [19].
FIGURA 3.21 Mantenimiento persistente de la secrecin de progesterona cclica, por administracin de
embriones de 14-16 das o de trofoblastina recombinante ovina (oTP): A. Mantenimiento de la secrecin de
progesterona despus de inyecciones diarias en el tero de una oveja cclica (T) comenzando desde el da 12
del ciclo, de un homogeneizado de embrin de 14-16 das (*)o de 21-23 das (o).
B. Mantenimiento de la secrecin de progesterona despus de la inyeccin diaria en el tero de una oveja
cclica (T) desde el da 10 y durante ocho das de oTP 340 g/da (). Control (o)[19].
El conceptas puede tambin secretar prostaglandina E, que actuando sobre las pequeas
clulas luteales del cuerpo lteo, estimule la secrecin de progesterona y, de ese modo,
mantener la funcin del cuerpo lteo.
En algunas especies animales como el hurn, el perro y los marsupiales coincide, que el
cuerpo lteo del ciclo tiene una vida tan larga como la preez, por lo que en esas especies
es posible que no sea necesario el reconocimiento de la gestacin por la madre.
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
PERTURBACIONES FUNCIONALES DEL CICLO OVARICO
OBJETIVO
El estudiante estar en la capacidad de identificar correctamente cualquier alteracin de las funciones del ciclo ovrico y aplicara las medida correctivas se
gun la perturbacin que se presente.
a.- ANESTRO. Concepto. Clasificacin y consecuencias econmicas
1.
Anestro con cuerpo luteo funcional
Anestro fisiolgico de la vaca gestante
Anestro por persistencia de cuerpo luteo
Anestro post-mortalidad embrionria o fetal precoz
2.- Anestro sin cuerpo lteo funcional
o Anestro post-partum
o Anestro por stress durante el parto
o Anestro debido a debilidad o marcada perdida de peso
o Anestro por Quistes ovricos
3.- Ovulacin silenciosa o celo silente
4.- Ciclos Anovulatorios
5.- Degeneracin Quistica de los Ovarios
o Quistes foliculares
o Quistes luteales
o Quistes de cuerpo luteo
6.- Ninfomana
7.- Manejo Clnico de la Hembra Anestrica
8.- Sndrome de la vaca repetidora
9.- Prevencin del Anestro
Este acpite es extra- curricular por lo cual los estudiantes del cuarto ao de Medicina
Veterinaria y Zootecnia tienen que realizar la investigacin mediante revisin bibliografica
actualizada.
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
REFERENCIAS BIBLIOGRFICAS
1). Me DNALO, L: Endocrinologa veterinaria y reproduccin. Nueva Editorial
Interamericana Me Graw-Hill, Mxico D.F., 1991.
2). GREEN, J.D. and G.W. HARRIS: J. of Phisiology, 108: 359,1949.
3). DERIVAUX, J.: Reproduccin de los animales domsticos. Ed. Acribia, Zaragoza,
1976.
4). NALBANDOV, A.V.: Fisiologa de la reproduccin. Ed. Acribia, Zaragoza, 1969.
5). CALDANI, M.; A. CARATY; J. PELLETIER; J.E. THIERY and L.H. TILLET:
Pulstil relase and its control. En: Reproduction in mammals and man. (Eds.) C.
Thibault; M.C. Levasseur and R.H.F. Hunter, Ellipses, Paris, 1993.
6). BARAM, T. and Y. KOCH: Gonadotropin releasing hormone in the milk, Science,
198:300,1977.
7). COMBARNOUS, YVES: Gonadotropina: Structure-Synthesis-Functions. En:
Reproduction in mammals and man. (Eds.) C. Thibault; M.C. Levasseur and R.H.F.
Hunter, Ellipses, Paris, 1993.
8). SHAMS, D.; F. HOFERT; E. SHALLENBERGER; M. HARTL and H. KARG:
Pattern of luteininzing hormone (LH) andfollicle stimulating hormone (FSH) in
bovine blood plasma after inyection of a synthetic gonadotropin releasing hormone
(GnRH). Theriogenology, 1:137-151,1979.
9). ROTTEN, D.: Regulation of FSH synthesis and secretion. En: Reproduction in
mammals and man. (Eds.) C. Thibault; M. C. Levasseur and R.H.F. Hunter, Ellipses,
Paris, 1993.
10). DUBOIS, P: The hipothalamic-pituitary axis, embriological, morphological and
functional aspects. En: Reproduction in mammals and man. (Eds.) C. Thibault;
M.C. Levasseur and R.H.F. Hunter, Ellipses, Paris, 1993.
11). BOTELLA LLUSIA, J.: Tratado de ginecologa, t. I, Fisiologa Femenina, Ed,
Cientfico Mdica, 1966.
12). WALTERS, D.L.; D. SHAMS and E. SHALLEBERGER: Pulsatile secretion of
gonadotrophins, ovarian steroids and ovaran oxytocin during the luteal phase of the
oestrous cycle in the cow. J. feprod. Fertif., 71: 479, 1984.
13). BRITT, J.H.: Current concepts of folllculogenesis and endocrinology, Embryo.
Transfer., 3(4): 1-3, 1988.
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
14). HANSEL, W. y EX CONVEY: Physiology of the estrous cycle, J. of Amin.
SG/.,57suppl.2:404.1983.
15). WETTEMANN, R.R; H.D. HAFS; L.A. EDGERTON and L.V SWANSON:
Estradiol and progesterone in blood serum during the bovine estrous cycle. J. of
Anim.Sci., 34(6):1 020,1972.
16). MARTAL, J. and L. CEDARD: Endocrine functions of the placenta. En
Reproduction in mammals and man. (Eds.) C. Thibault; M.C. Levasseur y R.H.F.
Hunter. Ellipses, Pars, 1993.
17). NISWENDER, G.D.; J.L. JUENGEL and W.J. Me GUIRE: Luteal function: The
estrous cycle and early pregnancy. Blol. feprod., 50:239,1994.
18). SHORT, R.V.: Role of hormones in sex cycles. En: Austin y Short, reproduction in
mammals. 3 hormones in reproduction, University Press, Cambridge, 1975.
19). LEYMARIE, R and J. MARTAL: The corpus luteum from cycle to gestation. En:
Reproduction in mammals and man. (Eds.) C. Thibault; M.C. Levasseur and R.H.F.
Hunter, Ellipses, Pars, 1993.
20). HANSEL, W. En: Ffeproduction in domestic animals. H.H. Col y RT. Cupps., vol.
1, Academic Press. 1959.
21). MALTIER, J.R; C. LEGRAND and M. BREUILLER: Parturition. En:
feproduction in mammals and man. (Eds.) C. Thibault; M.C. Levasseur and R.H.F.
Hunter, Ellipses, Pars, 1993.
22). SHELDRICK, E.L. and A.RF. FLINT: Regression of the corpora ltea in sneep In
responso to cloprostenol in not affected by loss of luteal oxytocin after
histerectomy. J. feprod. Frtil., 68: 155, 1983.
23). SATO, E.; TE. ISHIBASHI and A. IRITAN: Purification and action sites of a
follicle stimulating hormone inhibitor from bovine follicular fluid. J. of Anim. Sci-,
55: 873-877,1982.
24). HENDERSON, K.M.; R FRANGHIMONT; M.J. LECONTEYERNA; N. HUDSON
and K. BALL Increase in ovulation rate after active inmunization of sheep with
inhibin partially purified from bovine follicularfluid. J. Endocr., 10:305-309,1984.
25). BOTELLA LLUSIA, J.: Las protenas placentarias hormonales y no hormonales,
En: La Placenta, fisiologa y patologa. Ediciones Daz de Santos, Madrid, 1993.
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
26). DADOUNE, J.R and A. DEMOULIN: Structure and functions of the testis. En:
Reproduction in mammals and man. (Eds.) C. Thibault; M.C. Levasseur y R.H.F.
Hunter, Ellipses, Paris, 1993.
27). SHEMESH, M.; W. HANSEL and KRW. CONCANNON: Testosterone synthesis in
the bovlne corpus luteum. Biol. Reprod., 13:490-493,1975.
28). WISE, T.H.; D. CATN; W.W. THACHER; A. RAMI LEHRER and M.J. FELOS:
Androstenedione, dehydroepiandrosterone and testosterone in ovaran vein plasma
and androstenedione in peripheral arterial plasma during the bovine oestrous cylce.
J. fteprod. Frtil., 66: 513-518, 1982.
29). PETERS, A.R.: Hormonal control of the bovine oestrus cycle. The natural cycle.
British Veterinary Journal, 141: 564, 1985.
30). CROSS, B.A.: The Hypotalamus. En: Reproduction in mammals. 3 hormones in
reproduction. Cambridge, University Press, 1975.
31). HARRISON, F.A.; R.B. HEAP; E.W. HoRTONea/.: Identification of prostaglandin
F2a in uterine fluid from the non-pregnant sheep with an autotrasplanted ovary. J.
of Endocrinology, 53: 215- 222,1972.
32). SHORT, R.V.: Species differences. En: Austin y Short, reproduction in mammals
4. Reproductiva Patterns. Cambridge University Press. 1975.
33). ARMSTRONG, D.T. and W. HANSEL: Alteration of the bovine estrous cycle with
oxytocin, J. of DairySci., 42: 533-542,1959.
34). GINTHER, O.J.; C.O. WODY; S. MAHAJAN; K. JANAKIRAMAN and L.E.
CASIDA: Effect of oxytocin administration on the oestrous cycle of unilaterally
histerectomized heifers. J. Reprod. Frtil., 14: 225, 1967.
35). Me CRACKEN, J.A.; J.C. CARLSON; M.E. GLEW et al.: Prostaglandin F2ot
identified as a luteolytic hormone in the sheep. Nature, 238:129,1972.
36). KNICKERBOCKER, U.; M. C. WILTBANK and G.D. NISWENDER: Mechanism
of luteolysis in domestic livestock. Domest. Anim. Endocrcnol., 5: 91, 1988.
37). FLINT, A.PF.; El. SHELDRICK; T.J. Me CANN and D.S.C. JONES: Luteal
oxytocin and PGF2a secretion at luteolysis in rumiants, Dom. Anim. Endocr.,
7:111-124,1990.
38). AULETTA, F.J. and A.RF. FLJNT: Mechanims controlling corpus luteum function
in sheep, cows, non human primates and women, especially in relation to the time of
luteolysis. Endocr. Rev., 9:88-106,1988.
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
39). STEVENSON, J.S.: CImical reproductive phisiology of the cow. En: Current
therapy in large animal terogenology. (Ed.) Robert S. Youngquist, Ed. W. B.
Saunders Company, Philadelphia, 1996.
40). KINDAHL, H.; J.O. LINDELL and L.E. EDQUIST: Relase of prostaglandin Fa
during the oestrus cycle. Acta Vet. Scand., suppl. 77; 143,1981.
41). GINTHER, O.J.: Local versus systemic utero-ovarian relationships in farm
animals. Acta Vet. Scand., suppl. 77:103, 1981.
42). KUDLAC, E. a L. HOLY: Rizeni a kontrola reprodukce ve veikochovech skotv.
Statni zemedeiske Nakiadatelstvi, Praha, 1984.
43). PHARRIS, B.D.; J.C. CORNETTE and G.D. GUTKNEHT: Vascular control of
luteal steroidogenesis. J. eprod. Frtil., suppl. 10: 97, 1970.
44). NISWENDER, G.D. and T.M. NETT: The corpus luteum and its control. En: The
physioiogy of reproduction. (Eds.) E. Knobil; J. Neill et al., Raven Press, Ltd, New
York, 1988,
45). BAIRD, D.T: Reproductive hormone En: Reproduction in mammals. Hormones in
reproduction, University Press, Cambridge, 1975.
46). MOOR, R.M. and LEA ROWSON: Local mamtenance ofthe corpus luteum in
seheep with embryos transferred to various isolated portions of the uterus.J.
Reprod. Frtil-, 12:539-550, 1966.
47). HAFEZ, E.S.E.: Reproduccin e inseminacin artificial en animales. 5ta. ed. Nueva
Editorial interamericana Mac Graw-Hill, Mxico D.F, 1989.
48). POLGE, C.; R.M. MOOR; B.N. DAY; D.W. BOOTH and L.E.A. ROWSON:
Embryo numbers and lteal maintenance durng early pregnancy in swine. J.
Anim.Sci, 26(6): 1499,1967.
49). ZARCO, L; G.H. SSTABENFELDT; S. Bosu el al.: Modification of F2a sntesis
and relase in the ewe during the initial establishment of pregnancy J. Reprod.
Frtil., 83:527-536,1988.
50). MARTAL,J.;M.C. LACRoix;C.LouoEs,C.efa/.: Trophoblastin. an antiluteolytic
protein present in early pregnancy in sheep. J. Reprod. Frtil., 56:63-73,1979.
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
4. FECUNDACIN Y GESTACIN
La maduracin, la liberacin y la fecundacin de los vulos, son procesos esenciales en la
reproduccin animal, previos a la gestacin y consiguiente creacin de un nuevo ser. Una
vez que el vulo es expulsado del folculo, es necesario que sea fecundado por un
nemaspermo para que contine el proceso de multiplicacin. Con la fecundacin del vulo
se inicia el desarrollo de un nuevo individuo que durante toda la etapa intrauterina
depender de su madre.
El logro de la conjugacin de ambos gametos depende fundamentalmente de su integridad
y del momento en que se realice la unin sexual o la inseminacin artificial, por lo que se
debe conocer el momento en que se produce la ovulacin en cada especie, la supervivencia
de sus gametos y el tiempo que dura su trnsito a travs de las vas genitales de la hembra.
Viabilidad de las clulas germinales
El conocimiento de la supervivencia y la conservacin del poder fecundante de los
espermatozoides en el tracto genital femenino, es un elemento de gran importancia
prctica. Se sabe que estos pierden su capacidad fecundante, antes que su motilidad. De
acuerdo con los clculos que se han realizado y con los resultados de la reproduccin en el
ganado bovino, se considera que el perodo mximo de supervivencia de los
espermatozoides para obtener una fertilidad ptima, es de 24 h, aunque su supervivencia
con baja fertilidad puede llegar a 48 h o algo ms [1].
Birgitta Larson [2] recobr solo un pequeo nmero de espermatozoides mviles en el
oviducto de novillas 24 h despus de la inseminacin. En la yegua, sin embargo, los
espermatozoides pueden conservar su motilidad y fertilidad, hasta 5 das, y en la perra
estos se han recuperado, en el lumen uterino, 11 das despus del coito [3].
La supervivencia de los ovocitos es menor que la de los espermatozoides, En la mayora de
las especies se considera que son capaces de fertilizacin y desarrollo hasta 12-24 h
despus de la ovulacin. La perra constituye una excepcin, al tener una supervivencia de
sus ovocitos mucho mayor en el oviducto, donde pueden vivir de 4-8 das, Probablemente
esto sea debido a que en esta especie la expulsin del primer corpsculo polar puede
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
retardarse algunos das despus de la ovulacin, ya que ovulan ovocitos primarios que
maduran en el oviducto. Los ovocitos al llegar al istmo disminuyen muy rpido su
fertilidad y la pierden por completo al llegar al tero.
Sustancias inhibidoras de la motilidad de los espermatozoides
El plasma seminal presenta sustancias inhibidoras de la motilidad de los espermatozoides.
En los toros contiene una protena secretada por las glndulas anexas que fijan los iones
Ca2 necesarios a la motilidad. Los iones Zn++ secretados por la prstata bloquean la
actividad fosfolipasa A2 de la membrana citoplasmtica del espermatozoide til a la
formacin de productos desestabilizadores de la membrana como la fosfatidilcolina [4].
El esperma pasa desde el plasma seminal a los fluidos del tracto genital de la hembra y se
eliminan las sustancias inhibidoras. Esto ocurre en los animales de eyaculacin vaginal, en
el crvix y en los de eyaculacin uterina directamente en el tero. Los fluidos uterinos y
tubricos tienen dos funciones independientes: supervivencia y capacitacin de los
espermatozoides.
Transporte del esperma en el aparato genital femenino
En todas las especies de animales domsticos, as como en la humana, un nmero limitado
de espermatozoides alcanza el oviducto en menos de 15min [5].
Van Dermark y Moeller [6] demostraron que en la vaca esto sucede en 2-4 min
independientemente de que los espermatozoides estuviesen vivos o muertos y de que se
hubiera realizado la monta natural o la inseminacin artificial. Con posterioridad se
comprob que los espermatozoides vivos llegan al oviducto en mayor proporcin que los
muertos [2].
Independientemente del sitio donde se produzca la deposicin del semen, se establece una
prolongada y duradera corriente de espermatozoides hacia la vagina.
En las especies de eyaculacin de tipo vaginal, como la bovina, la ovina, la caprina y la
humana, el semen eyaculado en el coito es fuertemente rociado, sobre el orificio vaginal
del cuello uterino, por lo que se forma una laguna en el fondo de la vagina, pero no hay
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
evidencia de que durante el mismo el fluido seminal pase al tero. En las especies de
eyaculacin de tipo uterino, como la equina y la porcina, el semen es depositado en su
mayor parte en el tero; y en la canina, el semen es llevado al tero a travs del crvix
relajado por la actividad muscular del tracto genital [5].
Despus del coito los espermatozoides penetran en las micelas del moco cervical que les
proporciona proteccin y nutricin y donde son transportados por el canal cervical, en su
transporte intervienen la motilidad espermtica, y fundamentalmente las contracciones del
miometrio y mesosalpinx [7].
Las micelas del moco cervical ayudan a los espermatozoides en su ascenso por el conducto
cervical, donde muchos son atrapados en los complejos pliegues de las criptas cervicales,
en las que se forma un reservorio de ellos, que queda protegido del ambiente hostil de la
vagina y de la fagocitosis, ya que en las secreciones cervicales hay pocos leucocitos, en
comparacin con las de la vagina o el tero. A la vez el crvix filtra los muertos y
defectuosos y, posiblemente, participe en su capacitacin. Cuanto mayor sea la cantidad de
espermatozoides que entren al reservorio cervical, mayor ser el nmero de ellos que
lleguen al oviducto y, por tanto, aumentarn las probabilidades de fecundacin [8].
Al penetrar los espermatozoides en el tero, no encuentran un medio favorable, debido a
que su presencia all estimula la invasin de leucocitos que los fagocitan [5]. La
infiltracin de leucocitos en la luz uterina y su activacin para ingerir espermatozoides
despus del apareamiento, parece ser el mecanismo ms importante para eliminarlos. No es
necesario que estn muertos o daados para que sean fagocitados [8].
Durante un experimento, fueron recobrados ms espermatozoides de la unin terotubrica que del oviducto, 2 h despus de la inseminacin artificial, mientras que a las 12 y
a las 24 h sucedi todo lo contrario, lo que indica que dicha unin tero-tu banca acta
como otro reservorio o barrera, desde aqu los espermatozoides pueden ser liberados al
oviducto [2]. Ms espermatozoides fueron recobrados del oviducto izquierdo que del
derecho, desproporcin que no estuvo asociada a la actividad ovrica.
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
Los espermatozoides que logran entrar al istmo tubrico, se dirigen a la ampolla donde se
encontrarn con el vulo. No se sabe si el esperma se transporta a la ampolla por una
relajacin momentnea de la unin ampulostmica o por su propia motilidad. Dentro del
oviducto se ha observado una marcada disminucin de espermatozoides en la ampolla [9].
Es posible que los que no estn asociados con el huevo sean considerados como perdidos y
sean transportados fuera del oviducto [2].
Fenmeno de la capacitacin
Austin [10] calific como fenmeno de la capacitacin a la necesidad que tienen los
espermatozoides de determinadas especies de animales de experimentar algunos cambios,
para lo que tienen que permanecer un tiempo en el aparato genital de la hembra antes de
tener la capacidad de fecundar. Hasta ahora se ha demostrado la necesidad de capacitacin
de los espermatozoides del toro, el verraco, el gato, el conejo y la rata [11].
Los espermatozoides del conejo pueden ser capacitados en el tero de las perras [12], y los
del toro en las vas genitales de la oveja y la cerda [13].
La capacitacin puede representar un proceso selectivo para evitar la fertilizacin por
espermatozoides dbiles o anormales, puesto que implica su supervivencia durante varias
horas en el tracto genital de las hembras [14]. Este proceso, debe comprender la
destruccin o la eliminacin de un factor presente en el plasma seminal, debido a que la
incubacin en plasma seminal de espermatozoides ya capacitados motiva la prdida de su
capacidad de fecundar. El factor de descapacitacin parece no ser especfico de las especies
[11].
Reaccin acrosmica
La reaccin acrosmica (RA) es un evento que comprende fusiones mltiples entre la
membrana plasmtica de la esperma y la membrana acrosmica externa, lo que trae como
resultado la formacin de una membrana vesicular hbrida. El contenido acrosomal
hidrolizado por las enzimas del acrosoma, es liberado a travs de la membrana vesicular
fenestrada (Figura 4.1). Las vesculas son eliminadas del esperma antes de que penetre la
zona pelcida y que la membrana acrosomal interna expuesta acte de manera directa con
dicha zona [15].
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
Resultados de mltiples investigaciones evidencian que en los mamferos el esperma
fertilizante sufre la reaccin acrosmica en la superficie de la zona pelcida [15].
Cambios caractersticos de la motilidad durante la capacitacin
Los espermatozoides maduros progresan linealmente, es el llamado movimiento
progresivo. Estudios microcinematogrficos han puesto de manifiesto un desplazamiento
lateral de sus cabezas: la ligera asimetra en el movimiento de las colas induce una
progresin ligeramente curvada, con cambios regulares de derecha a izquierda y viceversa.
Ello es debido a una rotacin de 180 de las cabezas en un plano simtrico lo que da la
impresin de una progresin en lnea recta. Durante la capacitacin se producen dos
cambios [4]:
1. Se incrementa la amplitud del batimiento de las colas a causa de su mayor flexibilidad
y, principalmente, de la pieza intermedia; como consecuencia la cabeza se balancea
ligeramente en oposicin a la cola (movimiento de ltigo).
2. Cesa la rotacin de la cabeza de modo que la progresin se hace global-mente circular
(Figura 4.2).
La capacitacin y la hiperactivacin pueden ser experimentalmente disociadas. Esto indica
que diferentes mecanismos y estructuras estn involucrados [16].
FIGURA 4.1 Reaccin acrosmica: A. Acrosoma intacto en el espermatozoide antes de la reaccin
acrosmica (RA). B. La fusin mltiple de la membrana plasmtica (MP) y la membrana acrosmica externa
(MAE) lleva a la formacin de vesculas quedando la membrana fenestrada, esta permite la salida de
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
contenido acrosomal hidrolizado. C. Despus de la RA las vesculas son eliminadas del espermatozoide y es
expuesta la membrana acrosmica interna (MAI) el segmento ecuatorial permanece intacto [16]
FIGURA 4.2 Cambios de la motilidad del espermatozoide durante la capacitacin [4]
Fertilizacin
La fertilizacin del vulo en las especies de animales domsticos, tiene lugar en la ampolla
tubrica. Para que esta se produzca, es esencial que arribe all cierto nmero de
expermatozoides dotados de capacidad fecundante. En los mamferos superiores la
fertilizacin del ovocito es monoespermtica. Contribuye a ello el nmero relativamente
pequeo de espermatozoides que alcanza la ampolla (Tabla 4.1) y muy especialmente la
rpida reaccin de la zona pelcida, cuando un espermatozoide logra penetrarla, la que
bloquea la entrada del resto, bloqueo de la polispermia [3].
La polispermia (participacin de ms de un espermatozoide en la fertilizacin) es
patolgica en los mamferos y casi siempre produce la muerte del embrin, ya que origina
la formacin de proncleos supernumerarios incompatibles con el desarrollo normal del
embrin. En estos casos el nmero de cromosomas es triplolde (3 n) o mayor an,
alteracin de carcter letal que, por lo general, no les permite vivir ms all de la
implantacin [17].
La zona pelcida constituye, tambin, una barrera eficaz que impide la fertilizacin
heterogmica. La digestin enzimtica de esta zona permite la fertilizacin interespecies in
vitro [11].
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
TABLA 4.1 NMERO DE ESPERMATOZOIDES PRESENTES EN EL ITSMO Y
LA AMPOLLA POCAS HORAS ANTES DE LA OVULACIN
Especie
Itsmo
Ampolla
No. Espermatozoides/ ovocito
Vaca
2000-4000
> 10
10
Oveja
400-1 600
17-25
15
Cerda
6-10000
1 100
130
El huevo llega al conducto tubrico rodeado por el cmulo ooforo y la corona radiada,
cuyas clulas foliculares en los rumiantes y en la yegua se dispersan rpidamente en la
ampolla del oviducto, lo que permite que los espermatozoides entren en contacto directo
con la membrana pelcida del ovocito.
Los espermatozoides contienen en el acrosoma una enzima, la hialuronidasa y, adems,
factores dispersores activos como la neuraminidasa y enzimas tipo tripsina, como la
acrosina [14] capaces de despolimerizar al cido hialurnico del cumulas. La actividad
proteoltica es esencial para la penetracin de la zona pelcida, que parece ser la envoltura
ms difcil de penetrar.
En algunas hembras (mujer, ratona, cobaya) la penetracin del espermatozoide, en el
ovocito es completa, en otras est limitada a la cabeza, y la cola queda en el exterior, como
sucede en la vaca, la oveja, la perra y la coneja [17]. Cuando el espermatozoide penetra en
el espacio perivitelino, las membranas plasmticas del vulo y el espermatozoide se funden
y sus materiales citoplasmticos se mezclan.
Los proncleos masculino y femenino se forman de 11-39 h despus de la fecundacin en
la vaca y se pueden diferenciar porque el femenino es asimtrico. Despus de formarse los
proncleos, sus cromosomas se unen para originar el cigoto. La unin de los cromosomas
es la singamia, con la que concluye la fertilizacin.
Los principales resultados de la fertilizacin son [18]:
1. La restauracin del nmero diploide de cromosomas.
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
2. La determinacin del sexo del cigoto.
3. La iniciacin del clivaje.
Clivaje
Con la divisin del cigoto en dos blastmeras da inicio el clivaje, serie di rpidas
divisiones celulares mitticas, que conducen a la produccin de un nmero
progresivamente mayor de clulas sucesivamente ms pequeas pero que conservan el
nmero diploide de cromosomas (2n). Aunque durar te el clivaje hay un aumento del
nmero de blastmeras, no hay aumento di volumen protoplasmtico, sino por el contrario,
disminuye este como resultado de la actividad metablica. El clivaje termina al iniciarse la
etapa c blastocisto y transcurre en un trmino de 7-8 das en la vaca, de 4-5 en yegua, de 57 en la oveja y de 5-6 en la cerda.
Migracin del huevo
En muchas especies de animales domsticos, despus de la fertilizacin el huevo
permanece en el rea mpulo-stmica por 3 o 4 das, luego pasa rpidamente a travs del
istmo y la unin tero-tubrica al tero (Figura 4.3): esta travesa, por lo general, se realiza
en el estado de mrula o principio del blastocisto.
En la cerda puede pasar ms rpido (2o 3 das), mientras que en la perra puede requerir de
6-8 das y en la gata de 5-9 das. La duracin del trnsito es independiente del tamao del
animal, del largo del conducto tubrico y de la duracin de la gestacin [17].
Bajo condiciones normales el huevo permanece en el oviducto hasta que se forma el
cuerpo lteo, y el endometrio bajo la influencia de la progesterona le puede proporcionar el
medio y la nutricin adecuada [5]; su llegada en el momento apropiado es esencial, ya que
si esta es prematura o si se retarda ocasiona la muerte del embrin.
En la yegua solo los huevos fertilizados pasan al tero. Los que no lo han sido quedan
retenidos en el oviducto, donde sufren degeneracin y fragmentacin [11].
La implantacin del blastocisto en el tero no tiene lugar tan pronto llega all por un
perodo de tiempo el embrin permanece libre en el lumen uterino; su implantacin en la
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
vaca ocurre a los 20 o 30 das, en la yegua alrededor de las siete semanas, en la oveja de
11-14 das, en la cerda de 4-7 das y en la perra y la gata cerca de una semana [19].
Los requerimientos nutritivos del embrin al inicio son cubiertos por: el material que trae
el huevo, las secreciones tubricas y las sustancias en las secreciones uterinas. Despus de
la implantacin, el embrin es mantenido principalmente a travs de la placenta [19],
La migracin del huevo est asegurada por la corriente de las secreciones tubricas hacia el
tero, condicionadas estas por los movimientos de los cilios vibrtiles del epitelio y por las
contracciones peristlticas de la musculatura tubrica. En la ampolla parece que son ms
importantes los movimientos ciliares, mientras que el trnsito a travs del istmo depende
esencialmente de las contracciones peristlticas de la musculatura tubrica [17].
FIGURA 4.3 Migracin del huevo y sus estadios.
Se ha demostrado que el vulo queda retenido en el oviducto a consecuencia de un bloqueo
temporal del conducto. Esta contraccin muscular depende de la accin estrognica. La
progesterona, por el contrario posee un efecto depresor sobre la actividad muscular de la
ampolla, lo cual lleva consigo el relajamiento de la musculatura, un ensanchamiento de la
luz tubular y una aceleracin en el descenso de los vulos [17].
Cuando el blastocisto est libre en el tero, se mueve, presumiblemente en relacin con las
contracciones de las paredes uterinas, las que pueden resultar en la migracin del huevo
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
hacia el cuerno opuesto, fenmeno muy frecuente en la oveja, la yegua, la cerda, la perra y
la gata, pero raro en la vaca [5].
Nutricin hemotrfica
Cuando el blastocisto aumenta de tamao le resulta insuficiente la nutricin embriotrfica,
mediante la que se nutra por difusin de la leche uterina. Entonces se produce la
implantacin y con esta comienza la organizacin de las membranas placentarias, las que
le permitirn el acceso al sistema circulatorio materno; de este modo se opera la transicin
a la nutricin hemotrfica.
Membranas fetales
Las membranas fetales o extraembrionarias son rganos auxiliares que funcionan durante
la vida fetal, para la nutricin, la proteccin y la excrecin de anhdrido carbnico,
nitrgeno y otros productos finales del metabolismo, hasta que el nuevo ser en desarrollo
pueda tener una vida independiente, fuera del claustro materno. Estas membranas son:
corion, amnios, saco vitelino y alantoides.
El corion, el saco vitelino y la alantoides, sirven para la nutricin del embrin, y el amnios
lo protege de la desecacin y de la accin de los agentes externos. El corion recubre todo el
proceso embrionario y desarrolla en su superficie las denominadas vellosidades crlales,
que establecern la placenta fetal.
El saco amnitico se halla inmediatamente alrededor del embrin, dentro del corion. Se
encuentra lleno de lquido que ofrece proteccin a los suaves tejidos del embrin y que
aumenta su volumen con ms intensidad durante el cuarto y el quinto mes de la gestacin.
Al final de esta el lquido contiene gran cantidad de moco denso, con un color blanco
semitransparente. Este moco se supone que proceda de la saliva y las secreciones
nasofarngeas del feto, el que durante la gestacin se cree traga lquido amnitico y expulsa
saliva.
El feto normal no inhala lquido amnitico a los pulmones [5]. Al concluir el perodo de la
gestacin el volumen total de fluido amnitico en la vaca es de 4-5 L y en los pequeos
rumiantes no sobrepasa los 500 ml. Una caracterstica fundamental de dicho lquido,
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
especialmente en el ganado vacuno, es que en el parto lubrica el canal obsttrico y facilita
la expulsin del feto. La presencia de meconio en este, por lo general, es sntoma de asfixia
fetal.
En la superficie interna del amnios de la vaca se encuentran unas pequeas formaciones,
ms perceptibles del tercer al sptimo mes de la gestacin, las que han sido llamadas placas
glicognicas [20] o placas amniticas [5], cuyo significado no est debidamente aclarado.
Estas placas se encuentran en menor grado en yeguas, ovejas y cabras, pero no estn
presentes en el amnios de la cerda, ni en el de los carnvoros.
El saco vitelino es una estructura primitiva que se establece tempranamente en los
mamferos. Despus de un corto perodo desaparece en los rumiantes y los suidos, pero
persiste de 4-6 semanas en los quidos. La mayor importancia del saco vitelino, es que a su
nivel y a expensa del mesodermo, que forma parte de su pared, se originan los vasos
sanguneos vitelinos. El saco vitelino se va atrofiando y se reduce de tamao, tomando la
forma de una pequea bolsa pediculada unida al intestino medio. Esta formacin se llama
finalmente vescula umbilical [21].
La alantodes penetra a travs del ombligo corporal y ocupa la cavidad del celoma
extraembrionario, queda cubierta con la capa mesodrmica y se extiende entre el amnios y
el corion [22]. Se liga ntimamente al corion para formar el alantocoron y al amnios para
crear el alantoamnios. A partir del mesodermo alantoideo se desarrollan vasos sanguneos
que se extienden hacia el corion a travs del llamado pedculo alantoideo [21].
La alantoides est completamente formada a los 24 o 28 das de la gestacin y se encuentra
lista para participar en la funcin placentaria a los 40 o 60 das [23]. En las vellosidades del
alantocorion se ramifican los capilares sanguneos, los que al unirse componen dos arterias
y una o dos venas umbilicales. Este sistema vascular se designa con el nombre de
circulacin alantoidea [3]. Las arterias se conectan con la aorta y la vena con el hgado y
con la vena cava posterior del feto [5],
La cavidad de la alantoides est llena del lquido alantoideo, acuoso y ms oscuro que el
lquido amnitico, contiene albmina, fructosa y urea, provenientes de productos de
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
desecho de los riones fetales, los que pasan a esta desde la vejiga a travs del cordn
umbilical por intermedio del uraco.
A mediados de la gestacin tiene el mismo volumen que el amnitico, pero luego aumenta
hasta alcanzar al final de esta de 8-15 L en la vaca, de 8-18 L en la yegua y de 0,5-1,5 L en
la oveja y la cabra; de 100-200 M en la cerda, de 10-50 M en la perra y de 3-15 en la gata
[5],
En el lquido alantoideo se puede encontrar, flotando, uno o ms cuerpos esponjosos; estos
son suaves, de color amarillento, de forma irregular y del tamao de la palma de la mano.
Se designan estos cuerpos romanees y se supone que son residuos de la leche uterina.
Cuerpos similares se hallan tambin en la oveja (otomanes), cabra (capramanes) y equinos
(hipomanes) [22].
En la vaca desde los 60 a los 90 das de la gestacin es frecuente observar en el
alantocorion, precipitaciones de calcio, en forma de bandas, las que ms tarde desaparecen
al depositarse este mineral en los huesos. Depsitos similares han sido reportados en cerdas
entre 20 y 46 das de la gestacin [5].
En los equinos y los carnvoros el amnios, fijado solo por el cordn umbilical, flota en el
lquido alantoideo, pero en los rumiantes y los suidos se establece una estrecha fusin entre
el alantoamnios y el alantocorion, en la zona prxima al dorso del feto. formando el
amniocorion. El lquido alantoideo queda, portante, en estas especies contenido
principalmente en los extremos de la alantoides, uno de cuyos extremos se encuentra en el
cuerno no grvido. En los bvidos, segn Arthur [23] al final de la gestacin, la presin del
lquido alantoideo acumulado tiende a separar de nuevo al alantoamnios del alantocorion,
pudiendo llegar a semejar la disposicin de los sacos alantoideos y amniticos que tienen
los equinos y los carnvoros. En la vaca y en la cerda la alantoides no llega a los extremos
del corion, por lo que estos privados de irrigacin sangunea sufren necrosis.
Cordn umbilical
El cordn umbilical est constituido por dos arterias, dos venas que se renen para
conformar una sola, el conducto vitelino atrofiado y el uraco casi siempre obliterado en el
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
momento del parto, unidos por la gelatina de Wharton y cubiertos en el ternero y el cerdito
en toda su extensin por el amnios. En el potro su porcin distal est cubierta por la vaina
que establece la continuidad de la alantoides.
A travs del cordn umbilical se lleva a cabo toda la comunicacin entre la madre y el feto.
El del ternero mide de 30-45 cm de largo y el del cerdito unos 25 cm, mientras que el del
potro tiene casi 1 m de largo.
Placenta y placentacin
La placenta est constituida por dos partes, una fetal, el alantocorion , con sus vellosidades
y otra maternal, el endometrio con anfractuosidades par; recibir las vellosidades. La ntima
unin que se establece entre ambos componentes de la placenta se designa como
placentacin y a travs de esta e feto recibe los elementos nutritivos indispensables a su
desarrollo y elimine los productos de desasimilacin, adems de cumplir otras tareas de
defensa (barrera placentaria) y de sntesis de hormonas y enzimas.
Clasificacin de las placentas
Las placentas de los mamferos superiores se pueden dividir en completas o verdaderas
(vera o coniucta), en las que las vellosidades profundizan dentro del endometrio, por lo que
la placenta se expulsa con porciones de tejido uterino, llamado de decidua (que cae); y en
semiplacentas (apposita), en las que las vellosidades coriales no sobrepasan la capa
epitelial del endometrio, hay solo una yuxtaposicin del epitelio corinico sobre el
endometrial [21]. En ellas debido a la ligera unin de las placentas (fetal y maternal), en el
parto la fetal se expulsa sola, sin restos de placenta maternal.
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
NMERO DE CAPAS DE TEJIDO
De acuerdo con el nmero de capas de tejido que separan la sangre fetal de la maternal, las
placentas se pueden clasificar en: epiteliocorial, sindesmocorial, endoteliocorial y
hemocorial.
Epiteliocorial. Se conservan todas las capas que separan la sangre fetal de la maternal, es
decir, endotelio de los capilares sanguneos, tejido conectivo y epitelio del alantocorion;
endotelio de los capilares sanguneos, tejido conectivo y epitelio del endometrio. Tienen
este tipo de placenta la yegua, la burra y la cerda.
Sindesmocorial. En este tipo de placenta desaparece el epitelio del corion y quedan cinco
capas, pero se considera como semiplacenta y est presente en los rumiantes. En la vaca se
ha observado que el epitelio de las carnculas uterinas, se va perdiendo a partir del sptimo
mes de la gestacin y que en los casos en que este desaparece completamente, hay
retencin placentaria [24].
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
Endoteliocorial. En esta placenta verdadera o completa, hay una mayor penetracin de las
vellosidades coriales, que causan gran erosin del endometrio, con destruccin del epitelio
y tejido conectivo, quedando la lmina sincitiotrofoblstica en contacto directo con el
endotelio de los capilares sanguneos, por lo que en su expulsin es difcil separar la parte
fetal de la maternal [21]. Esta es la placenta tpica de los carnvoros.
Hemocorial. Es una placenta verdadera, propia de los primates y los roedores. En ella hay
gran destruccin del endometrio y se establecen espacios intervellosos, que se convierten
en verdaderas lagunas sanguneas a niveles profundos del endometrio. Las vellosidades
coriales se baan ntegramente en la sangre materna. En el parto la placenta se desprende
ntegra y queda una capa germinativa de mucosa excenta de erosin, que regenera toda la
mucosa del endometrica. Cuando se expulsa esta placenta, se presenta una hemorragia
profusa [21].
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
DISTRIBUCIN DE LAS VELLOSIDADES CURIALES
Las placentas tambin se clasifican de acuerdo con la distribucin de las vellosidades
coriales, las semiplacentas pueden ser difusas o cotiledonadas, y las placentas verdaderas,
zonales o discoidales.
Difusas. En estas planenlas las vellosidades se encuentran distribuidas de manera
uniforme por toda la superficie del alantocorion, como sucede en la yegua, la cerda, la
camella y el tapir. La cerda tiene la particularidad de presentar dos pequeas zonas carentes
de vellosidades (manchas calvas) situadas irregularmente [14].
Cotiledonadas. Las vellosidades se agrupan en zonas circunscritas de la superficie del
corion denominadas cotiledones. A estos cotiledones corresponden engrasamientos de la
mucosa uterina, las carnculas. La unin de los cotiledones con las carnculas constituyen
los placentomas. Este es el tipo de placenta caracterstico de los rumiantes. En la vaca, las
carnculas se encuentran en nmero de 100-120, dispuestas en cuatro hileras paralelas.
Durante la gestacin se hipertrofian y alcanzan las mayores unos 5 cm de altura y de 10-12
cm de longitud. Su superficie es convexa y su unin con el endometrio queda reducida a un
pedculo, por donde transitan los vasos que van a irrigar las criptas de las carnculas
(Figura 4.4).
Las carnculas de la oveja y la cabra naturalmente son ms pequeas. Las ovejas tienen de
60-80 con la superficie cncava, mientras que la superficie de las de la cabra es plana.
Zonales. Propias de los carnvoros; las vellosidades coriales se encuentran agrupadas en
una faja o anillo circular y el resto del corion es liso (laeve), es decir, carece de
vellosidades. Las placentas zonales tienen un hematoma marginal, con un pigmento verde
en las perras y pardo en las gatas.
Discoidales. Son propias de los primates, en ellas las vellosidades corinicas se encuentran
concentradas en forma de un disco circular u oval. Este tipo tambin lo tienen los roedores
y el oso, teniendo en estos la particularidad de tener un hematoma marginal con un
pigmento pardo.
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
FIGURA 4.4 Placentoma de la vaca [22]
Fusin de las placentas en gestaciones mltiples
En todos los casos de gestaciones dobles en vacas, en un mismo cuerno se encontr fusin
de los vasos del alantocorion de ambos fetos. En le casos en que la gestacin de cada feto
se desarroll en un cuerno distinta la anastomosis vascular se observ en el 67 % de los
casos [25]. La parabiois que se establece ocasiona que los fetos hembras de las gestaciones
con el feto macho, sean freemartin. Esta forma especial de intersexualidad es consecuencia
del eritromosaiquismo que se produce en esos casos.
En las gestaciones dobles de vidos y cpridos se fusionan los corion pero no las
alantoides, que son las que conducen los vasos fetales a placenta. En las gestaciones
mltiples de los suidos no hay fusin de la placentas, quedando cada una independiente
(Figura 4.5).
Se han reportado casos de freemartinismo en ovejas, cabras y cerdas, pe parece que solo se
ha demostrado debidamente su presencia en la oveja [26]
Circulacin sangunea fetal
La circulacin sangunea fetal es sumamente interesante, porque durante desarrollo fetal la
respiracin es placentaria y tan pronto nace el animal: convierte en una respiracin
pulmonar.
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
En el feto es necesario que una buena parte de su sangre venosa, media las arterias
umbilicales pase por la placenta, donde debe oxigenarse y eliminar los desechos
metablicos, para transformarse en sangre arterial, Igualmente se requiere que al pulmn
solo le llegue la sangre necesaria para nutricin, ya que en la etapa fetal dicho rgano no
funciona, adems el tema circulatorio, al producirse el nacimiento, tiene que estar en
disposicin de cambiar bruscamente la respiracin placentaria por la pulmonar.
Para poder cumplir esas funciones y afrontar esos cambios, el feto al nacer dispone de las
arterias y las venas umbilicales, del agujero de Botal o foramen oval y del conducto
arterioso de Botal o ductus arteriosus y en rumiantes, los roedores, los primates y los
carnvoros, tambin del conducto venoso o conducto de Arancio [27].
FIGURA 4.5 Fusin del corion y de la alantoides en embriones de la vaca (A); fusin del corion en los
embriones de oveja y cabra (B) y no se fusionan ni el corion ni la alantoides en la cerda (C).
Durante la vida fetal, sangre procedente de las arterias iliacas, llega a la placenta a travs
de las arterias umbilicales y se transforma en sangre arterial. De all en la yegua y en la
cerda es conducida por la vena umbilical a la red capilar del hgado, en las especies que
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
tienen el mencionado conducto de Arando, parte de la sangre de la vena umbilical es
transportada seguidamente a la vena cava posterior por medio de este. A medida que
avanza la gestacin la cantidad de sangre que va de manera directa al hgado es cada vez
mayor mientras que la que va por el conducto de Arancio es cada vez menor.
Este conducto se origina en un seno venoso del interior del hgado, por la confluencia de
las venas porta y umbilical [28] y se dirige directamente a la vena cava posterior. De este
modo llega a la aurcula derecha la vena cava posterior con sangre arterial procedente de la
vena umbilical, mezclada con su sangre venosa, ms la sangre venosa de la vena porta, y
adems la vena cava anterior con sangre tambin venosa (Figura 4.6).
Para limitar la cantidad de sangre que pase al pulmn procedente del ventrculo derecho a
la que se requiere para cubrir sus necesidades nutricionales, el feto dispone del agujero
oval, orificio existente en el tabique interauricular que est provisto de una vlvula, la
vlvula de Eustaquio. Esta, muy desarrollada en el corazn fetal, desva la corriente
sangunea como si fuera un dique, que permite que parte de la sangre de la aurcula
derecha pase a la izquierda y evita que refluya a la derecha.
El resto de la sangre de la aurcula derecha pasa al ventrculo derecho y sale por la arteria
pulmonar, donde sufre otro desvo antes de llegar al pulmn, a travs del conducto
arterioso, que comunica a la arteria pulmonar con la aorta posterior. Este conducto tiene un
calibre mayor que las ramas de la arteria pulmonar que van a los pulmones. Se cree que la
sangre transportada a la aurcula derecha por la vena cava posterior, pasa en gran parte, si
no toda, a la aurcula izquierda mediante el agujero oval, mientras que la que llega por la
vena cava anterior pasa al ventrculo derecho [28].
Al nacer el ternero y terminar con ello su vida fetal, cambia inmediatamente su respiracin
placentaria, dependiente de la madre, a la pulmonar. La desviacin de la corriente
sangunea, aplica la vlvula de Eustaquio contra el agujero de Botal, lo que produce su
cierre y queda como una profunda fosa oval en la aurcula derecha.
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
La aspiracin de sangre por el pulmn, al respirar, provoca que el conducto arterioso se
colapse, con lo que queda establecida la circulacin menor 10 pulmonar. El conducto
arterioso se transforma entonces en una cuerda fibrosa, el ligamento arterioso [28].
FIGURA 4.6 Circulacin fetal [27]
Ms tarde las arterias umbilicales, que han perdido su funcin, se transforman en los
ligamentos redondos de la vejiga urinaria. En los rumiantes y los carnvoros la vena
umbilical y el conducto de Arancio se atrofian y se convierten en el ligamento redondo del
hgado.
La sangre del feto y la de la madre circulan independientes en conductos completamente
separados, y todos los intercambios entre ellas se producen por difusin [29]. Las dos
circulaciones estn lo suficientemente relacionadas en la unin del corion y el endometrio,
para que el oxgeno y los nutrientes pasen de la sangre maternal a la fetal y se eliminen
productos de desecho, en direccin contraria [30].
tero grvido
Despus de la fecundacin siguen desarrollndose y profundizndose las transformaciones
endometriales para poder asegurar la alimentacin y la supervivencia del nuevo individuo.
Las glndulas uterinas producen una rica secrecin, pero tambin otras partes del tero
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
(miometrio, vasos sanguneos) evolucionan para adaptarse a las necesidades de recibir el
conceptus.
La evolucin del tero se encuentra condicionada por la hipertrofia masiva de las clulas
musculares, que pueden aumentar su tamao hasta siete veces. Esta evolucin se debe en
parte a la presencia de los estrgenos y en parte a la del conceptus.
Durante este proceso el tero aumenta de peso hasta 10 veces en relacin con su peso
original; en los primeros dos meses el aumento del tero se halla condicionado por el
aumento de los lquidos fetales y desde el quinto mes en adelante por el tamao del feto.
La pared se torna ms fina y alcanza de 2-3 mm de espesor (4,6 y 10 mm cuando el tero
est vaco, no gestante), tambin, los ligamentos anchos uterinos aumentan su volumen
hasta seis veces, los vasos sanguneos sufren considerables modificaciones adaptndose a
las necesidades del conceptas.
El tero en su evolucin cambia de posicin en el transcurso de la preez, saliendo de la
cavidad pelviana y entrando en la abdominal. En la mayora de los casos cae en la bolsa
omental, que lo fija para evitar un gran movimiento en el abdomen; as en la mayora de
los casos el tero fijado en la bolsa omental ocupa la parte derecha del abdomen y la panza
la parte izquierda solo en algunos casos el tero preado puede estar fuera de la bolsa
omental y entonces se mueve libremente en el abdomen.
Posicin y movimientos del feto durante la gestacin
En las primeras etapas de la gestacin, las circunstancia de espacio son favorables al feto,
por lo que su posicin es variable. Cuando la gestacin progresa su eje longitudinal se hace
paralelo a la direccin longitudinal del tero y ms tarde adquiere una flexin mxima de
las extremidades y la cabeza, para ocupar el menor espacio posible dentro del tero.
Antiguamente se crea que la posicin que adoptaban todas las especies en la etapa final de
la gestacin era la inferior o ventral y que durante la fase de dilatacin se produca su
rotacin para adoptar la posicin superior. Hoy da se sabe que esto es vlido para la yegua
y la perra, pero no para la vaca y la oveja, especies en la que solamente se produce en la
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
fase de dilatacin el cambio de actitud de la flexin a la extensin, ya que en la ltima
etapa de la gestacin adquieren la posicin superior.
En el cuarto mes de la gestacin se notan en la vaca movimientos activos del feto que
reciben el nombre de pisciformes, los que son manifiestos cuando palpamos el tero por
exploracin rectal. A partir del quinto mes se pueden apreciar movimientos del feto en el
flanco inferior derecho, si se dan a la pared abdominal pequeos movimientos con el puo.
Nmero de fetos en el tero
En los animales domsticos, ciertas influencias de la domesticidad pueden aumentar la
disposicin para la multiparidad, que parece hereditaria, como por ejemplo, es abundante
pienso y el modo de criarlos con vistas a un madurez precoz (oveja). La enanizacin
(perra) reduce el nmero de cras.
Por cada 100 partos, la yegua tiene por trmino medio uno gemelar, ms de la mitad de sus
preeces gemelares no llegan a trmino. La frecuencia de partos gemelares en la vaca es de
un 2 % (razas europeas) y solo de un 0,5 % en las razas ceb. Las ovejas primparas tienen
partos mltiples con menos frecuencia que las multparas.
La cerda es ms frtil en las razas ordinarias (9-12 cras) que en las ms refinadas (6-9
cras), partos de 12-16 son muy raros.
Las perras presentan oscilaciones considerables en sus partos, segn sus razas e individuos;
las grandes razas tienen, por lo general, de 7 -10 cras; las perras de tamao mediano
suelen tener de 6-8 cras, como mximo diez; las de razas enanas tienen como mximo
cuatro cras. Los partos con una sola cra se presentan en todas las razas y tamaos, pero
son ms frecuentes en las perras pequeas. La gata tiene de 3-5 cras, se han reportado
casos excepcionales con ocho cras.
Duracin de la gestacin
La duracin de la preez es influida por una serie de factores de los cuales el papel ms
importante lo desempaan, sobre todo, los factores genticos y los ambientales.
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
Los factores genticos incluyen la raza (Tabla 4.2), el genotipo del feto y la influencia de
los padres, especialmente la paterna. Entre los ambientales se encuentran las condiciones
internas, como por ejemplo, las relaciones hormonales feto-madre, el tamao y el sexo del
feto, la edad de la madre, etc. El sexo del feto influye en la duracin de la preez, tanto en
ganado de carne como en el de leche, el feto macho prolonga la gestacin de 1 -2 das,
tambin la edad de la madre influye; en caso de vacas viejas la gestacin es ms
prolongada.
TABLA 4.2 DURACIN DE LA GESTACIN EN DIFERENTES RAZAS DEL
GANADO BOVINO
Raza
Ganado lechero
Promedio (das)
Mnimo (das)
Mximo (das)
Brown Swiss
289.7
270
306
Ayrshire
277.9
Frisian
276.3
240
333
Guernsey
283.9
262
359
Holstein Fnsian
278.8
270
285
Jersey
278.8
Shorthorn de leche
281.7
Frisian de suecia
281.7
260
300
Aberdeen angus
279
Shorthorn de carne
282
273
294
285.1
243
316
285
Gsnado de carne
Hereford
Ceb
Hoy da se discute la dependencia de la duracin de la preez con la temporada del ao,
aunque en la yegua es mucho ms manifiesto. Se sabe que cuando las yeguas son gestadas
en el mes de diciembre la gestacin se prolonga un promedio de 10,4 das.
Para cada especie de los animales domsticos existe un promedio de duracin tpica de la
preez, excluyendo las variaciones externas (Tabla 4.3).
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
TABLA 4.3 DURACIN DE LA GESTACIN EN DIFERENTES ESPECIES
DOMSTICAS
Especie
Vaca
Duracin promedio (das)
282
Yegua
333
Oveja
150
Cabra
150
Cerda
115
Perra
63
Gata
58
Coneja
30-32
La madre durante la gestacin
En la preez se producen una serie de reacciones sinrgicas y antagnicas entre el feto y la
madre en absoluta armona fisiolgica; el equilibrio recproco se establece por las
secreciones endocrinas en colaboracin con el sistema nervioso.
La significacin de la madre para el feto lgicamente es extraordinaria, no solo durante el
desarrollo intrauterino, sino tambin en el puerperal.
La gestacin aunque es un estado fisiolgico impone a la madre determinadas exigencias,
por ello en el organismo materno se desarrollan algunos cambios tpicos que influyen en
todas las funciones corporales.
Celo en la preez
La inhibicin de la funcin estral est presente en la mayora de las hembras domsticas y
el celo no se presenta hasta que la cra no es destetada y se reanuda el ciclo estral, sin
embargo, un determinado nmero de vacas (5 % o ms) puede aparecer en celo en el
primer tercio de la gestacin.
Aunque el ganado vacuno es un donador o productor muy pobre de hormonas estrognicas,
durante la preez, sobre todo desde el cuarto mes de la gestacin, aumenta el tenor de
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
estrgenos, siendo la fuente de ellos los ovarios y la placenta; estos posibilitan el aumento
de la mitosis y el crecimiento celular. La sntesis de la protena contrctil del miometrio
(actiomiosina) estimula la permeabilidad vascular del tero y aumenta la sensibilidad del
miometrio.
Metabolismo
Sin contar que con la gestacin recesa la actividad estral, la preez cambia en la madre
especficamente el metabolismo general. Adems del aumento del apetito, se mejora y se
intensifica la digestin, se aprovechan bien los nutrientes (anabolismo de la gestacin) y
los animales mejoran su estado fsico, por lo menos en la primera fase de la preez.
En la segunda mitad de la gestacin el feto se desarrolla ms rpidamente y sus exigencias
aumentan en especial en minerales y vitaminas, por lo que es necesario dirigir la
alimentacin no solo cuantitativa sino tambin cualitativamente.
Respiracin y circulacin
Durante la preez avanzada el organismo materno, se encuentra sobrecargado,
especialmente los aparatos respiratorio y circulatorio. En algunos individuos se aprecian
edemas hasta ascitis. Desde el comienzo de la segunda mitad de la preez, en los
mamferos multparos, la respiracin se hace ms costal a consecuencia de la disminucin
de la actividad del diafragma, producida por la presin del tero. Los animales en estado
avanzado de la preez respiran con dificultad y ms de prisa.
Para aumentar el espacio torcico, los carnvoros adoptan la posicin de perro sentado, o
intentan reducir la presin de los rganos abdominales sobre el diafragma echndose de
lado.
Alimentacin durante la gestacin
Teniendo en cuenta las demandas de nutrientes del feto, la composicin de la dieta de las
hembras gestantes debe ser variada y dietticamente sin defectos; en esta etapa hay que
eliminar todos los alimentos que pudieran producir cualquier alteracin (timpanismos,
diarreas, etc.) que amenacen al nuevo ser.
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
En la tabla 4.4 aparecen las necesidades de elementos minerales y vitamina A en la vaca
gestante.
TABLA 4.4 NECESIDADES DE MINERALES Y VITAMINA A EN LA VACA
GESTANTE
Elemento
Ca
Durante la preez
60-85 g
60-65 g
1 00-200 mg
Na
24-32 g
Vitamina A
134 000 UI
Diagnstico de la gestacin
El diagnstico de la gestacin en las especies mayores tiene gran importancia econmica y
es un elemento imprescindible en el control de la reproduccin. En la vaca el diagnstico
se puede realizar por mtodos clnicos o de laboratorio. Los primeros comprenden,
mtodos de exploracin externa e interna.
Mtodos de exploracin externa
Los mtodos de exploracin externa tienen poca aplicacin en las especies de animales
mayores, ya que los de exploracin interna permiten en estas especies un diagnstico
mucho ms temprano y muy certero. Sin embargo, cuando se van a secar las vacas, a los
siete meses de gestacin, es conveniente comprobar su preez de forma sencilla y segura
mediante la palpacin externa. Esto es muy conveniente porque se puede haber incurrido
en un error diagnstico o se puede haber producido un aborto que no fue observado.
La exploracin externa comprende:
1. Inspeccin
2. Palpacin
3. Auscultacin,
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
INSPECCIN
Por inspeccin es posible apreciar el aumento de volumen y el cambio de forma del
abdomen, pero debido al gran tamao de la panza estos cambios, por lo general, son de
escaso valor durante la primera mitad de la gestacin. A los siete meses si el especialista se
para detrs del animal, el flanco izquierdo a nivel del muslo es convexo por la presencia de
la panza, y el derecho cae verticalmente en la porcin superior, mientras que se le aumenta
el volumen en la parte inferior, por la presencia del feto [22]. No obstante, esto es solo un
signo probable de gestacin.
Por inspeccin tambin se pueden percibir, a partir de los siete meses de la gestacin,
movimientos del feto en forma de sacudidas y de elevaciones de la pared abdominal
derecha, por delante de la cabilla, que aparecen y desaparecen [27]. La observacin de
estos movimientos tiene gran inters porque asegura que el feto est vivo, aunque el no
apreciarlos no quiere decir lo contrario.
PALPACIN
A partir del sexto mes de la gestacin, al imprimir pequeos movimientos al feto, mediante
presiones rtmicas ejercidas en el flanco derecho de la vaca, es posible palpar al feto que
choca contra la pared abdominal, percibindose como un cuerpo duro. Para ello el
operador apoya su mano izquierda en el ngulo externo del rea, y con el puo derecho da
movimientos bruscos a la pared abdominal, por delante de la babilla derecha, en un rea de
aproximadamente 30 cm y espera que el feto baloteado venga a chocar contra la pared, en
la que lo puede palpar.
La palpacin de un cuerpo duro que viene a chocar es garanta de gestacin, pero en
algunas ocasiones puede suceder que no se palpe, si la vaca est hoy gorda o si el tero
fuera del omento se ha situado debajo de la panza.
AUSCULTACIN
La auscultacin del abdomen en la vaca, llevada a cabo para percibir los latidos cardacos
fetales, confronta la dificultad del gran nmero de ruidos, que resultan de la actividad de su
aparato digestivo, por lo que el no escucharlos no dice que no exista una gestacin, ni que
el feto haya muerto.
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
La poca ms propicia para escuchar estos latidos es a partir del sptimo mes de la
gestacin, en la planicie del ijar derecho, aplicando el estetoscopio o el odo a la altura de
la articulacin fmoro-tibio-rotuliana. Su gran frecuencia (120-150 latidos/ min) permite
diferenciarlos de los de la madre. La percepcin de los latidos cardacos fetales asegura el
diagnstico de la gestacin, as como la vitalidad del feto.
Mtodos de exploracin interna
La gestacin en la vaca se puede determinar con un alto grado de precisin, mediante la
exploracin por va rectal. La palpacin rectal del tero y de su contenido, es el mtodo de
eleccin para calcular el tiempo de la gestacin.
Los elementos que se pueden utilizar para el diagnstico de la gestacin durante la
exploracin rectal son:
1. Presencia de un cuerpo lteo en el ovario.
2. Palpacin de la vescula amnitica.
3. Fluctuacin en el cuerno por presencia de lquido.
4. Palpacin de los vasos de la alantoides.
5. Palpacin de la membrana corioalantoidea (sensacin de doble pared).
6. Asimetra de los cuernos.
7. Palpacin del feto.
8. Presencia de placentomas.
9. Hipertrofia y frmito de la arteria uterina media.
De estos, son signos precoces inequvocos de la gestacin, la palpacin del la vescula
amnitica, la de los vasos de la alantoides, la de la membranal corioalantoidea, la del feto y
la de los placentomas, y a partir del quinto mes la hipertrofia y el frmito de la arteria
uterina media. Por el contrario constituye un signo inequvoco de que no hay gestacin, la
inexistencia de cuerpo lteo v n los ovarios.
Para obtener buenos resultados con la exploracin rectal, los tactores ms importantes son:
estar familiarizado con las particularidades anatmicas del tero normal, no grvido, y
realizar un examen cuidadoso y metdico.
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
DIAGNOSTICO PRECOZ DE LA GESTACIN
El signo que ms precozmente permite el diagnstico de la gestacin en la vaca, mediante
la palpacin por va rectal, es la presencia o la ausencia del cuerpo lteo de gravidez.
Durante el perodo comprendido entre 21 y 24 das posteriores a haber recibido un
servicio, el cuerpo lteo habr regresado si no ha habido gestacin. La ausencia de un
cuerpo lteo funcional en esos momentos, indica con un 100 % de efectividad que la vaca
no est gestante, mientras que su presencia permite diagnosticar la gestacin con un 90 %
de seguridad.
El primer signo inequvoco de gestacin que se puede apreciar por exploracin rectal, es la
presencia de la vescula amnitica, la que se puede palpar desde la cuarta a la sexta semana
de la gestacin, gracias a su turgencia en ese perodo, como una estructura ovoidal, que
flota en el lquido alantoideo [31].
Se encuentra situada en la porcin libre el cuerno grvido, cuerno con ms fluido y pared
ms fina, ipsilateral al ovario que tiene el cuerpo lteo de gestacin. A las cuatro semanas
la vescula tiene aproximadamente 2 cm de dimetro y algo ms de 3 cm en la sexta
semana.
La palpacin de la vescula amnitica se realiza, al deslizar la porcin libre del cuerno
presuntamente grvido, entre el pulgar y el ndice o entre el pulgar y el resto de los dedos.
La palpacin de esta vescula no debe ser utilizada como medio rutinario de diagnstico,
por el peligro de producir algunos casos de abortos. Su uso se justifica en casos en que sea
necesario establecer un diagnstico de gestacin precozmente.
Otro signo inequvoco de gestacin que se puede percibir, en el citado perodo precoz de la
gestacin, es la presencia de los vasos de la alantoides. El alantocorion que durante los
primeros das sufre una elongacin, el da 35 de la gestacin tiene una longitud aproximada
de 40 cm, ocupando todo el cuerno grvido, se va llenando progresivamente del lquido
alantoideo y forma un gran saco alantocorinico que va distendiendo el cuerno.
A todo lo largo de este saco alantocorinico marchan los vasos de la alantoides, sobre una
delgada lmina de tejido conjuntivo, los que se pueden percibir en este perodo, al frotar
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
las paredes del cuerno entre el pulgar y el ndice, dando la sensacin de la existencia de un
cordn fino dentro del cuerno grvido. La palpacin de estos vasos no debe tampoco ser
utilizada como diagnstico de rutina, por el peligro de provocar abortos. Su palpacin trae
como consecuencia hemorragias, segn se ha observado en animales sacrificados despus
del diagnstico.
Los dos mtodos de diagnstico precoz descritos deben ser precedidos por la palpacin de
los cuernos en toda su extensin. Si estos se hallan vacos tendrn consistencia de carne
[31]. Si hay gestacin estarn presentes fluidos en uno o en ambos cuernos, lo que resulta
en una fluctuacin caracterstica. La existencia de lquido en el cuerno se puede comprobar
por percusin; segn Abelein, se coloca el pulgar en contacto con la pared del cuerno y se
percute con el ndice en la pared opuesta del cuerno contra el pulgar.
El tratar de diagnosticar la presencia de los vasos de la alantoides sin previamente
comprobar la existencia de fluidos en el cuerno, conduce en algunas ocasiones a errores
diagnsticos, al confundir otras estructuras con dichos vasos [31].
Aunque se puede pensar que el diagnstico de la vescula amnitica puede ocasionar ms
abortos que la palpacin de los vasos de la alantoides, las experiencias realizadas [32]
mostraron una tendencia a que se produjera un nmero mayor de abortos en los animales
que se sometieron a la palpacin de los vasos alantoideos (Tabla 4.5).
TABLA 4.5 PRDIDAS FETALES DESPUS DEL DIAGNSTICO PRECOZ DE
LA GESTACIN [31]
Palpador
Fluctuacin
Mtodo de exploraci
n empleado
Fluctuacin
Fluctuacin ms
Totales
Palpador A N
solamente
122
ms amnios
106
vasos alantoides
107
335
Prdidas (%)
2 (1.6)
2 (3.8)
8 (7.5)
14 (4.2)
Palpador B N
75
91
99
265
Prdidas (%)
4 (5.3)
5 (5.5)
8 (8.1)
17 (6.4)
Palpador C N
79
126
87
292
Prdidas (%)
5 (6.3)
11 (8.7)
11 (12.6)
27 (9.2)
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
DIAGNSTICO DE LA GESTACIN A PARTIR DE LOS 60 DAS
En el diagnstico de la gestacin a partir de los 60 das tienen importancia los elementos
siguientes:
1. Asimetra de los cuernos.
2. Palpacin de la membrana corioalantoidea.
3. Palpacin del feto.
4. Presencia de placentomas.
5. Hipertrofia y frmito de la arteria uterina.
6. Examen vaginal.
Asimetra de los cuernos
El tero sano no grvido muestra una simetra casi completa. En las vacas, el cuerno que
estuvo gestante es ligeramente mayor, como consecuencia de la hiperplasia que siempre
acompaa a las hipertrofias. En las novillas la simetra de los cuernos es completa. Desde
la sexta semana de la gestacin (en las novillas incluso antes) se produce una asimetra
uterina cada vez ms notable, con adelgazamiento de la pared del cuerno, el que a las 10
semanas alcanza el grosor de un brazo.
La asimetra de los cuernos se debe valorar a nivel de la bifurcacin de los cuernos, para
ello se coloca el dedo del medio o corazn en la bifurcacin de los cuernos y se hace la
retraccin del tero hacia el interior de la cavidad pelviana y entonces con los dedos se
palpan las porciones libres de los cuernos y se comparan, uno con el otro.
Desde el final del primer mes aparece la fluctuacin del tero, la que se hace ms notable
en la medida que van aumentando los lquidos fetales, los que le dan al tero una
consistencia suave y elstica. El cuerno no grvido, que solo recibe una porcin del saco
corioalantoideo queda atrasado en cuanto a su desarrollo, adoptando el tero forma similar
a un guante de boxeo.
Palpacin de la membrana corioalantoidea
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
La membrana corioalantoidea solo est unida al cuerno en sus cotiledones, por lo que si se
agarra suavemente una porcin del cuerno y se permite que se deslicen entre el pulgar y el
ndice, la membrana corioalantoidea primero y la pared uterina despus, se siente la
sensacin de presencia de una doble pared, signo este caracterstico de que existe una
gestacin y que resulta, adems, muy til en su diagnstico diferencial con algunas
patologas que pueden semejar una gestacin, en las que falta este signo (pimetra,
hidrmetra, momificacin fetal, maceracin fetal). A las 10 semanas de la gestacin con
frecuencia es fcil detectar la gravidez por palpacin de dicha doble pared en el cuerno no
grvido.(Figura 4.7).
Palpacin del feto
El feto no puede ser palpado hasta que la vescula amnitica haya perdido su turgencia y se
encuentre flcida, lo que ocurre alrededor de la novena semana de gestacin. En ese
momento es posible palpar el feto de manera directa en el cuerno uterino, como una
formacin dura.
Posteriormente al aumentar cada vez ms el volumen del cuerno grvido, resulta imposible
palpar directamente el feto y es necesario lograr su palpacin mediante su peloteo. El
peloteo se practica al golpear suavemente el cuerno gestante 2 o 3 veces, con lo que se
imprime movimiento a los lquidos que contiene y se desita al feto, el que viene a chocar
contra la pared uterina, en la que se puede percibir con la palma de la mano apoyada en su
cara dorsal.
60 das de gestacin
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
FIGURA 4.7 Gestacin de siete semanas. Palpacin de la doble pared [22].
La palpacin del feto no solo permite el diagnstico de la gestacin, sino que constituye un
elemento valioso para diagnosticar su duracin. El tiempo de la gestacin se puede calcular
por la longitud que tenga el feto desde la nuca hasta el maslo de la cola, valindonos de los
estudios de sus dimensiones corporales por meses, realizados por algunos autores [27]
(Tabla 4.6).
TABLA 4.6 LONGITUD DEL FETO POR MESES
Tiempo de gestacin (meses)
1
Longitud (cm)
0,8
15
28
40
52
70
80
90
Medir la longitud del feto, por lo general, se hace difcil, por lo que puede ser sustituida
ventajosamente por la medida de la cabeza fetal, la que se puede hacer con mayor precisin
y resulta de especial valor para calcular el tiempo de la gestacin. La cabeza del feto se
localiza revisando delicadamente el cuerno que contiene ms fluido entre el pulgar y los
restantes dedos hasta encontrarla. Entonces se mide su longitud con la mayor precisin
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
posible, por comparacin con el ancho de los dedos. La manipulacin no tiene peligro
cuando se hace con suavidad [31] (Tabla 4.7).
TABLA 4.7 ESTIMADO DE DURACIN DE LA GESTACIN POR LA MEDIDA
DEL LARGO DE LA CABEZA DEL FETO
Duracin (das)
68-70
Medida (cm)
1,5
80
3,5
90
5,5
100
7,5
110
120
10,5
Presencia de placentomas
Aunque su formacin es ms temprana son palpables, como un grano de frjol, en la pared
del tero cerca de los dos meses y medio. Segn la gestacin avanza, la flacidez del tero y
su aumento de tamao facilitan su palpacin. En ocasiones se comete el error de confundir
dichos placentomas con los ovarios o viceversa.
La presencia de placentomas es un signo de gestacin, pero se debe tener presente que en
animales que han parido recientemente se palpan las carnculas en el tero.
Durante el quinto y el sexto mes de la gestacin el tero cae al suelo del abdomen y es
cubierto por asas intestinales, por lo que es muy frecuente que no se puedan palpar en ese
perodo. Tambin es posible que falten en el cuerno no grvido si este no particip en la
placentacin.
Hipertrofia y frmito de la arteria uterina
La arteria uterina nace, en la vaca, de la iliaca interna muy cerca de su origen en la aorta,
de un tronco comn con la umbilical (aunque en ocasiones se origina individualmente)
gana el ligamento ancho por el que desciende hasta la cara dorsolateral del tero,
inmediatamente por delante del cuerpo.
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
Cuando la preez progresa, el tero cae a la cavidad abdominal y la arteria es arrastrada
con el ligamento cranealmente, hasta que en la segunda mitad de la gestacin puede ser
localizada 5 o 10 cm por delante del cuerpo del ilion. La arteria uterina media no puede ser
confundida con la iliaca interna, que est firmemente unida por una fascia al cuerpo del
ilion, ya que ella por correr por el ligamento ancho, es movible, por lo que se puede
desplazar de 10-15 cm [5].
La arteria uterina suple la mayor parte de la sangre del tero, por lo que en la preez
aumenta progresivamente su calibre ante las crecientes demandas del feto. La palpacin de
los cambios que se producen en el tamao de esta arteria y en las caractersticas de su
pulso, son de valor diagnstico en la gestacin, especialmente durante el quinto y el sexto
mes, y un auxiliar en el diagnstico diferencial con el pimetra y el hidrmetra. De las tres
arterias que llegan al aparato genital es la nica interesante a la exploracin rectal [14].
En novillas desde los 60-75 das de la gestacin se puede notar un cambio en el dimetro
de la arteria uterina correspondiente al cuerno grvido, que puede ser de 0,15-0,3 cm. En
vacas viejas el cambio se puede notar a los 90 das cuando el dimetro es de 0,3-0,45 cm
[5].
La arteria a los seis meses de la gestacin es del grueso del dedo meique, a los siete meses
del grueso del dedo del medio o corazn y de 8-8 1/2 meses del grueso del pulgar [31]. La
arteria que corresponde al cuerno no grvido tambin aumenta su dimetro, pero su
aumento no es tan grande y se produce ms tardamente. Si ambas arterias estn
igualmente aumentadas, se debe sospechar una gestacin gemelar bicornual.
Como resultado de la hipertrofia de la arteria, sus paredes se adelgazan y el curso de la
sangre es ms rpido, por lo que sus ondas pulstiles dejan de ser un verdadero pulso y se
convierten en una especie de trepidacin o frmito. Este frmito se puede sentir alrededor
de los 80-120 das de la gestacin. Para percibirlo la arteria se debe tomar suavemente
entre los dedos; demasiada presin digital puede producir un falso frmito.
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
Examen vaginal
Los cambios que se producen en la vagina durante la gestacin pueden tener valor en su
diagnstico, pero solo de importancia secundaria [5]. La mucosa vaginal es plida y seca
como en el diestro y el orificio cervical es plido y cerrado. La secrecin de las glndulas
del cuello, gelatinosa y correosa, adhesiva, forma un tapn para sellar el canal cervical,
este moco carmelita claro alrededor del da 60 de la gestacin est cubriendo el orificio o
haciendo prominencia en el 60 o 70 % de los casos. En el resto est presente en el canal
cervical, pero no es visible al examen vaginal [23]:
Caractersticas de la gestacin por meses
Primer mes. Hay un cuerpo lteo de gestacin. El tero se encuentra en la cavidad
pelviana, contiene de 30-60 mL de lquido alantoideo, ms abundante en el cuerno grvido
y una vescula amnitica de aproximadamente 2 cm de dimetro, que por su turgencia no
permite palpar al pequeo embrin (0,8 cm de longitud), que contiene en su interior. Se
pueden palpar los vasos de la alantoides que se extienden a lo largo del cuerno grvido,
sobre una delgada lmina de tejido conjuntivo.
Segundo mes. El tero que se mantiene en la cavidad pelviana presenta una asimetra
notable de los cuernos uterinos, alcanzando el grvido de 6-9 cm de ancho. Hay
fluctuacin en el cuerno, por la presencia de 200-500 mL de lquidos alantoideo y
amnitico. La vescula amnitica de 6-8 cm de dimetro ha perdido su turgencia y permite
palpar en su interior un teto de 6-8 cm de longitud.
Tercer mes. Se inicia el descenso del tero a la cavidad abdominal. El cuerno grvido mide
de 10-13 cm de ancho, contiene aproximadamente 1 L de lquidos fetales y en sus paredes
se palpan los placentomas de 1-1,5 cm, Por deslizamiento entre los dedos pulgar e ndice
de las paredes uterinas se puede percibir la sensacin de doble pared. El feto de 13-17 cm
de longitud se puede pelotear en el cuerno. Se puede apreciar el frmito de la arteria
uterina correspondiente al cuerno grvido, la que tiene de 0,3-0,5 cm de dimetro.
Cuarto mes. El tero ha cado a la cavidad abdominal. El cuerno grvido mide de 13-18 cm
de ancho, en sus paredes se palpan placentomas de 1,5-2,5 cm, contiene de 2-3 L de
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
lquidos fetales y un feto de 22-32 cm de longitud, que se puede pelotear. Se palpa la doble
pared y el frmito de la arteria uterina, de 0,5-0,8 cm de dimetro.
Quinto mes. El tero en cavidad abdominal, contiene de 4-5 L de lquidos fetales y un feto
de 30-45 cm de longitud que usualmente no es palpable. El cuerno grvido de 18-23 cm de
ancho presenta placentomas de 2,5-4 cm y permite palpar la doble pared. En la arteria
uterina de 0,6-1 cm de dimetro se puede apreciar el frmito de marcada importancia en
este perodo de la gestacin, en que, por lo general no se palpa al feto, el que se encuentra
en la profundidad de la cavidad abdominal, cubierto por las asas intestinales.
Sexto mes. El tero en cavidad abdominal contiene de 4-7 L de lquidos fetales y un feto
de 40-60 cm de longitud que comienza a ser palpable de nuevo. Los placentomas miden de
4-5 cm y se palpa la doble pared. La arteria uterina de 0,9-1,3 cm de dimetro presenta
frmito.
Sptimo mes. El tero comienza su ascenso dentro de la cavidad abdominal, contiene
alrededor de 10 L de lquidos fetales y un feto de 55-65 cm de longitud que, por lo general
es palpable. Los placentomas miden de 5-7 cm y se palpa la doble pared. La artera uterina
de 1,3-1,5 cm de dimetro presenta frmito.
Octavo mes. El tero contina su ascenso en la cavidad abdominal, contiene alrededor de
12 L de lquidos fetales y un feto palpable, de 60-85 cm de longitud. Los placentomas
miden de 6-9 cm y se palpa la doble pared, la arteria uterina de 1,3-1,7 cm de dimetro
presenta un notable frmito.
Noveno mes. El tero que ha ascendido dentro de la cavidad abdominal, puede contener
hasta 20 L de lquidos fetales y un feto de 70-100 cm de longitud. Los placentomas
alcanzan de 8-12 cm y se palpa la sensacin de la doble pared del tero. La arteria uterina,
que puede alcanzar 2 cm de dimetro, presenta un frmito muy notable (Tabla 4.8).
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
TABLA 4.8 CARACTERSTICAS DE DISTINTOS MOMENTOS DE LA
GESTACIN DE LA VACA
Das de
la
gestacin
Dimetro
cuerno
(cm)
Cantidad
fluido
(mL)
30
2-4
30-60
Dimetro Longitud
vescula
feto (cm)
amnitica
(cm)
1
0.8
40
3-6
75-100
2.5-3
1.7-2.5
50
5-7
90-200
3.5-5
3.5-5.5
60
6-9
200-450
6-7.5
6-8
70
80
90
120
150
7-10
9-12
10-13
12.5-18
18-23
350-650
500-800
750-1400
2000-3500
4000-5000
7-10
8-13
13.17
22.32
30.45
Tamao
placentamas
(cm)
0.5-0.7
0.5-1
1-1.5
1.5-2.5
2.5-4
Dimetro
arteria
uterina
(cm)
0.3-0.5
0.5-0.8
0.6-1
Frmito
arteria
uterina
Situacin del tero
Cuello en
pelviana
Cuello en
pelviana.
Cuello en
pelviana
Cuello en
pelviana.
Inicio
Descenso
Descenso
Descenso
tero en
abdominal
+
+
+
+
Presencia
del feto
cavidad
cavidad
cavidad
cavidad
Por peloteo
Por peloteo
Por peloteo
Por peloteo
cavidad Usualmente
no palpable
TABLA 4.8 (CONTINUACIN)
Das de
la
gestacin
Dimetro
cuerno
(cm)
Cantidad
fluido (mL)
Dimetro
vescula
amnitica
(cm)
Longitud
feto (cm)
Tamao
placentama
(cm)
180
4000-7500
40-60
4-5
Dimetro
arteria
uterina
(cm)
0.9-1.3
210
240
270
6300-10000
8000-12000
12000-20000
55-75
60-85
70-100
5-7.5
6-9
8-12
1.3-1.5
1.3-1.7
1.5-1.9
Frmito
arteria
uterina
Situacin del
tero
Presencia
del feto
tero en cavidad
+
++
++
Ascenso del tero
Ascenso del tero
Ascenso del tero
Usualmente
no palpable
Palpable
Palpable
Palpable
Posibilidades de error diagnstico
En ocasiones la vejiga urinaria llena, puede ser confundida con el cuerno gestante. Se debe
tener en cuenta que esta es redonda (no tubular), no est unida al cuello uterino y no tiene
bifurcacin. En estos casos el tero se encuentra desituado y se puede hallar a la derecha y
debajo de la vejiga urinaria [31]. Tambin se puede confundir el rumen con partes fetales,
pero este tiene consistencia pastosa. Otras posibles fuentes de error son el pimetra, el
hidrmetra, la momificacin fetal, las tumoraciones leuccicas y otras.
Diagnstico de la gestacin en la yegua
En la mayora de las yeguas es posible diagnosticar la gestacin a partir del da 30. Unos
ovarios altos en la regin sublumbar son indicios de que no hay gestacin o de que esta es
precoz. La dificultad de alcanzar los ovarios en su posicin acostumbrada, es casi prueba
concluyente de gravidez [23].
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
Primer mes. A los 30 das el cuerno grvido est turgente, redondeado y duro, en contraste
con el cuerpo uterino y el cuerno opuesto, que se mantienen flcidos. En el tercio inferior
del cuerno, por lo general el derecho, se localiza una formacin con fluctuacin elstica de
3-5 cm de dimetro, parecida a un huevo pequeo de gallina, la vescula corioalantoidea.
En este momento el tono uterino es elevado, por lo que las manipulaciones provocan
contracciones del cuerno, lateral y medialmente al crecimiento, que facilitan su
reconocimiento [33]. Una porcin del cuerpo lteo es palpable durante unos das despus
de la ovulacin en la regin de la fosa ovulatoria, hasta que es cubierto por una densa
tnica fibrosa ovrica [14]. Un recto tenso puede interferir con el examen, hacindolo
difcil o imposible, otra dificultad es la que presentan las yeguas paridoras que fueron
fecundadas durante el posparto.
Segundo mes. A mediados del segundo mes (45 das) la vescula corioalantoidea comienza
a penetrar en el cuerpo del tero, mide aproximadamente 5 cm de dimetro y de 5-7 cm de
longitud. Al final de este mes la mitad de la vescula se localiza en el cuerpo del tero y
tiene de 8-10 cm de dimetro semejando una naranja. Su consistencia an es relativamente
blanda. Desde mediados de este mes hasta el cuarto, hay marcada actividad ovrica y
durante el desarrollo de mltiples folculos uno o ambos ovarios son temporalmente
mayores que en los calores y en algunos casos mucho mayores [23]. Las ovulaciones y los
folculos indehiscentes dan lugar en ese perodo a cuerpos lteos accesorios.
Tercer mes. Al tercer mes el conceptas est totalmente incluido en el cuerpo del tero, con
la forma de un meln de aproximadamente 23 cm de longitud y 14 cm de dimetro. El
tero ha comenzado a descender y el borde dorsal se encuentra a nivel del pubis y se puede
confundir con una vejiga urinaria parcialmente llena, pero mediante peloteo se puede
palpar el feto.
Cuarto mes. Los ligamentos anchos se hallan tensos por la traccin ejercida por el tero y
arrastran consigo a los ovarios. Los cuernos se encuentran flexionados hacia la lnea media
sobre una masa fluctuante, el cuerpo del tero, el que no ha completado su descenso
permite la palpacin del feto.
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
Quinto-sptimo mes. La distensin uterina aparentemente es menor porque el tero ha
descendido y se ha dirigido hacia adelante, lo que dificulta el diagnstico, aunque en
muchas yeguas por peloteo se puede palpar el feto. Los ligamentos estn ms tensos y el
desplazamiento de los ovarios hacia la lnea media aproxima uno al otro y los priva de
movilidad, pero ya no es fcil encontrarlos.
Sptimo mes al parto. El ascenso del tero grvido comienza casi a los siete meses de la
gestacin, como resultado del considerable crecimiento del feto. La palpacin de este en
dicho perodo no presenta dificultades. No obstante, en esta etapa es ms difcil precisar el
tiempo de la gestacin, que durante sus primeros estadios.
Migracin del conceptos
Aunque en el ovario izquierdo de la yegua se produce un nmero mayor de ovulaciones
que en el derecho, en el cuerno derecho se desarrollan ms gestaciones, lo que indica que
las migraciones del conceptus, que tienen lugar antes de su fijacin, entre los das 12 y 14,
son ms frecuentes en la direccin del cuerno izquierdo al derecho.
Otros mtodos de diagnstico
Ultrasonografa
Recientemente la ultrasonog rafia basada en el efecto Doppler ha adquirido notable
desarrollo. En la vaca y en la yegua, en las que se puede colocar el ransductor cerca del
tero por la va rectal, los embriones se pueden detectar desde los 25 das (14 das en la
yegua). En otras especies con embriones pequeos y en las que el transductor tiene que ser
aplicado externamente, la deteccin es hecha ms tarde (30 das en la oveja y cabra y 32 en
la cerda). Hoy da se puede determinar el nmero exacto de embriones en la oveja y en la
cabra despus de 40 o 50 das, pero en la cerda esto no es posible [34].
En la perra desde el da 19 se pueden detectar las vesculas amniticas por ecografa; el
tejido placentario se puede notar el da 23 y el latido del corazn fetal el da 25. Este
diagnstico es 100 % seguro despus del da 28 y entre el da 28 y el 35 el nmero de fetos
puede ser fcilmente determinado. Diferentes estructuras fetales son identificables por
ultrasonido desde el da 40 en adelante [35].
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
En general, la seguridad en el diagnstico es alta en las hembras gestantes y es baja para
las diagnosticadas no gestantes.
Mtodos hormonales
RADIOINMUNOANLISIS (RIA)
Despus del servicio las concentraciones de progesterona producidas por el cuerpo lteo
permanecen altas en las hembras preadas, mientras del da 21-24 estos niveles caen
(valores menores de 2 mg/ml) en las hembras que no resultaron fecundadas (Figura 4.8).
Estos pueden medirse por el mtodo de RA, en sangre o en leche, utilizando fase slida en
tubos [36]. Los resultados del diagnstico basados en los niveles de progesterona son muy
seguros (p < 5 %) para diagnosticar las hembras no gestantes, pero pueden haber errores en
el de hembras gestantes (hasta 20 %) motivados por variaciones en la duracin del ciclo
estral, mortalidad embrionaria y persistencia del cuerpo lteo, particularmente en yeguas y
cabras [34].
FIGURA 4.8 Diagnstico precoz de gestacin en vacas y ovejas, mediante determinaciones por RIA de los
niveles de progesterona en el plasma. Contenido de progesterona en ovejas y vacas no preadas (._.)
comparada con el de estas especies preadas [37].
El nivel de precisin es similar para muestras de sangre o de leche.
En los ltimos tiempos se han preparado Kits diagnsticos que permiten al criador medir,
l mismo, los niveles de progesterona en leche, pero los resultados de estos Kits
permanecen cuestionables [34].
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
PROTENA B ESPECFICA DE PREEZ
La protena B especfica de preez (PSPB) ha sido medida en sangre perifrica, En
rumiantes la PSPB se puede detectar despus de 25 das de preez. Este diagnstico
comienza a popularizarse.
ESTRGENOS
Los estrgenos producidos por la placenta y el feto, o por el feto, se pueden utilizar para
diagnosticar la preez en yeguas o el nmero de fetos en las ovejas y las cabras. Aplicada
durante la preez tarda, 100-110 das en la oveja, este diagnstico tiene un alto grado de
seguridad para animales preados (90%) pero con frecuencia baja para aquellos no
preados [34]. Se consideran preadas las ovejas con concentraciones de estrgeno en el
plasma mayores de 0,3 g/mL
Los niveles de PMSG, en la actualidad gonadotropina corinica equina (eCG), se pueden
medir desde el da 35 en yeguas preadas. Este mtodo fue el primer diagnstico hormonal
de preez en animales de cra, pero hoy da es menos usado.
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
B.- PATOLOGIAS DE LA GESTACIN
OBJETIVO.
El estudiante tendr la capacidad y la habilidad de determinar, valorar, diagnosticar,
curar las diferentes patologas de la gestacin y prevenirlas.
1.- Prolapso Vaginal
2.- Torsin Uterina
3.- Gravidez extrauterina primaria y secundaria
4.- Muerte fetal y modificaciones post-mortem
- Momificacin
- Maceracin
-Enfisema de la Putrefaccin
5.- Aborto
- Aborto no infeccioso
- Aborto infeccioso
-Medidas de control y profilaxis
6.- Parto prematuro
7.- Prolongacin Patolgica de la Gestacin
8.- Placentitas
9.- Trastornos metablicos relacionados con la gestacin
- Toxicosis dravdica de la oveja
Con este acpite los estudiantes realizaran investigacin extra-curricular mediante revisin
de bibliografa actualizada y tendrn que hacer la exposicin del tema designado y se
reforzara el contenido y dudas que tengan los estudiantes respecto a la temtica.
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
REFERENCIAS BIBLIOGRFICAS
1). SALISBURY, G.W. y N.L, VAN DEMARK: Fisiologa de la reproduccin e
inseminacin artificial de los bvidos, 1969.
2). BIRGITTA, L: Distribution of spermatozoa in the bovino genital tract after artificial
insemination. Sveriges Lantbruks Universitet, Uppsala, Sweden, 1980.
3). AUSTIN, C.R.: Fertilization and development of the egg. En: fieproduction in
domestic animis. 2nd ed., (Eds.) H.H. Col and RT. Cupps, Academic Press, New
York and London, 1969.
4). FOURNIER-DELPECH, S. and C. THIBAULT: Acquisition of sperm fertilizing
ability. Epididymal maturation, accesory glands and capacitation. En: Reproduction
in mammals and man. (Eds.) C. Thibault; M.C. Levasseur and R.H.F. Hunter, Ellipses,
Pars, 1993.
5). ROBERTS, S.J.: Veterinary obstetrics and genital diseases. 2nd ed., Edwards Brothers,
Inc. Ann. Arbor, Michigan, 1971.
6). VAN DERMARK, N.L. and A.N. MOELLER: Speed of spermatozoan transport in
reproductive tract of estrous cow. Am. J. Physiology, 165: 674-679, 1951.
7). VAN DEMARK, N.L. and R.L. HAYS: Uterine motility responses to mating. Am.
J. Phisiol., 170: 518-521,1952.
8). HAFEZ, E.S.E.: Reproduccin e inseminacin artificial en animales. 5ta ed., Nueva
Editorial Interamericana Me Graw-Hill, Mxico D.F., 1989.
9). THIBAULT, C.; M. GERARD et Y HEYMAN: Transporte et survie des
spermatozoides chez la vache. Proc. Insem., 26: 343-356,1973.
10). AUSTIN, C.R.: Capacitation of spermatozoa. Intern. J. ofFert., 12(1):25, 1967.
11). PINEDA, M.H.: La biologa del sexo. En: Endocrinologa veterinaria y
reproduccin. (Ed.) L.E. Me Donald, Nueva Editorial Interamericana Me Graw-Hill,
Mxico D.F., 1991.
12). WILLIAMS, W.L.; T.D. ABNEY; H.M. CHERNOFF; W.R. DUKELOW and M.C.
PINSKER: Biochemistry and Physiology of the De capacitation Factor, J. Reprod.
Frtil., suppl. 2; 11, 1967.
13). MARTAL, J.: Les biotechnologies de la reproduction anmale. En: Biotechnologic.
4ta. ed., Rene Surban, Editorial Technique y Documentation, Lavoisier, Paris, 1993.
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
14). MARTIN MARTIN, E.: Fisiopatologa de la reproduccin, con sus bases sinpticas.
Instituto Experimental de Ciruga y Reproduccin de la Universidad de Zaragoza,
Espaa, 1985.
15). CROZET, N.: Fertilization in vivo and in vitro. En: feproduction in mammals and
man. (Eds.) C. Thibault; M.C. Levasseur and R.H.F. Hunter, Ellipses, Paris, 1993.
16). IANAGIMACHI, R.: Mammalian fertilization. En: The phisiologyofreproduction,
(Eds.) E. Knobil and J. Neill, Raven Press Ltd., New York, 1988.
17). DERIVAUX, J.: Reproduccin de los animales domsticos. Ed. Acribia, Zaragoza,
1976.
18). HAMILTON, W.J.; J.D. BOY y H.W. MOSSMAN: Embriologa humana, 1963.
19). COL, H,H. and PT. CUPPS: Reproduction in domestic animis. 2nd. ed, Academic
Press, New York and London, 1969.
20). PRIBYL, h.: Veterinarni prodnictvi praha, 1954.
21). VEGA, G.: Resmenes del desarrollo anatmico de aves y mamferos, Instituto
Cubano del Libro, La Habana, 1974.
22). HOLY, L.: Biologa de la reproduccin bovina. Editorial Cientfico-Tcnica, La
Habana, 1987.
23). ARTHUR, G.H.; D.E. NOAKES and H. PEARSON: Veternary Reproduction and
Obstetrics (heriogenology). 6 th. ed., Ed. Bailliere Tindall, London, Philadelphia,
Toronto, 1989.
24). GRUNERT, E.; Etiology of relamed bovine placenta. En: Current therapy in
theriogenology. (Ed.) D.A. Morrow, W.B. Saunders Company, Philadelphia, Toronto,
1980.
25). WILLIAMS, G.; I. GORDON and J. EOWARS: Observations on the frecuency of
fused fetal circulations in twin bearing cattle. Brit. Vet. J., 119: 467, 1963.
26). ALEXANDER, G. and D. WILLIAMS: Ovine Freemartins. Nature, 201:1296,
1964.
27). BENESCH, F: Obstetricia y ginecologa veterinarias. Editorial Labor, SA, Madrid,
1963.
28). SISSON, S.: Anatoma de los animales domsticos. 2da. ed., revisada, Salvat Editores,
S.A, Barcelona, 1974.
29). AREY, L.B.: Anatoma del desarrollo, Edicin Revolucionara, Instituto Cubano del
Libro, La Habana, 1968.
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
30). JAINUDEEN, M.R. and E.S.E. HAFEZ: Gestation, prenatal physiology and
parturition. En: Reproduction in farm animis. (Ed.) E.S.E. Hafez, 4th. ed., Lea and
Febiger, Philadelphia, 1980.
31). BALL, L.: Pregnancy diagnosis in the cow. En; Current therapy in thenogenology.
(Ed.) D.A. Morrow, W.B. Saunders Company, Philadelphia, Toronto, 1980.
32). ABBITT, B.; L. BALL; G.R KITTS et al.: Effects of three methods of palpation form
pregnancy diagnosis per rectum on embrionic and fetal attrition in COWS.JAVMA,
173:973,1978.
33). ZEMJANIS, R.: Reproduccin animal. Diagnstico y tcnicas teraputicas. Ed.
Umusa-Wiley, S.A., Mxico, 1966.
34). CHEMINEAU, R; D. CHUPN; Y COGNIE and J. THIMONIER: Control of
reproduction in domestic mammals. En: Reproduction in mammals y man. (Eds.) C.
Thibault; M.C. Levasseur and R.H.F. Hunter, Ellipses, Pars, 1993.
35). CHAFFAUX, SI: Reproduction of the cat and the dog. En: Reproduction in
mammals and man. (Eds.) C. Thibault; M.C. Levasseur and R.H.F. Hunter, Ellipses,
Paris, 1993.
36). REIMERS, T.J.; R.D. SMITH and S.K. NEWMAN: Evaluation of reproductive
perfomance using milk progesterone assay. En Livestock Reproduction in latn
Amrica Proceeding of the final research coordenation meeting. Bogot, Faoliacea,
Vienna, 1990.
37). STABLENFELDT, G.H.; L.E. EDQUIST; H. KINDHAL et al.: Practicalimplications
of recent physiologic findings for reproductive efficiency in cows, mares, sows,
andewes. JAVMA, 172(6):667-675,1978.
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
5. EL PARTO
El parto es el proceso mediante el cual el tero grvido expulsa al feto y sus envolturas una
vez concluida la gestacin. El parto marca la terminacin de la preez, as como el
momento en el que el feto es capaz de independizarse y existir fuera del tero.
Mecanismo del parto
Al finalizar la gestacin se produce el parto como consecuencia de cambios estructurales
en la placenta, interrelacin de hormonas maternales y fetales y factores fsicos o
mecnicos [1]. Existen diferencias entre las especies en el control del parto, siendo estas
ms notables entre los primates superiores y los animales domsticos. En este proceso
intervienen muchos factores que trabajan juntos en forma integrada para causar el parto,
cuando uno de los factores falla, otros intervienen para compensar su falta.
Durante la gestacin se mantiene un nivel alto de progesterona en el plasma perifrico
maternal, procedente del cuerpo lteo de gestacin o de la placenta.
La progesterona ejerce la accin llamada bomba de sodio que absorbe este catin al
interior de las clulas. Para compensar esta entrada, un equivalente de potasio sale al
exterior de las mismas y se origina as un equilibrio de las diferencias sodio-potasio desde
dentro hacia fuera de las clulas. Con esto el potencial de membrana y la excitabilidad
celular se hacen menores, esta es la causa de que la progesterona seda la fibra muscular
uterina, La declinacin de esta hormona al final de la gestacin, permite que sean
manifiestas las acciones etimulatorias de los estrgenos y de la prostaglandina sobre las
contracciones uterinas.
La cada de los niveles altos de progesterona prepartum, puede ser abrupta y restringida a
pocos das antes que comiencen las contracciones activas del tero, como sucede en la
vaca y la cabra (Figura 5.1) o gradual y extendida sobre la segunda mitad de la gestacin,
como ocurre en la coneja y la rata. No obstante, en la mujer y en los pocos primates
superiores de que se dispone informacin, hay poca o ninguna declinacin de los niveles
de progesterona del plasma perifrico prepartum [2], pero en su tero a trmino hay una
importante disminucin de un 40 % en la relacin P4/E2 [3] (Figura 5.2).
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
En las ovejas no es indispensable una disminucin de la concentracin de progesterona en
el miometrio para que se produzca el parto. Pero el incremento de la incidencia de distocias
cuando los niveles de progesterona permanencen elevados indica, que una disminucin de
la concentracin de progesterona o bajos niveles son deseables para que se produzca un
parto normal [4].
En el inicio del parto se considera significativa la liberacin de hormonas de la corteza
adrenal del feto. En el feto normal hay un aumento de los corticoides fetales cuando el
parto se aproxima. En el plasma de fetos ovinos, 20 das antes del parto se encontraron 5
mg/mL de cortisol, los que se elevaron a alrededor de 70 mg/mL el da del parto.
La importancia de la presencia de la hipfisis fetal y de un activo eje-hipfisis- adrenal, en
el comienzo del parto, se ha comprobado en la oveja, la cabra, la yegua, la coneja, la cerda
y la vaca. La destruccin experimental del eje adrenal hipofisario en estos animales resulta
en la prolongacin de la gestacin. Otro tanto ocurre cuando sufren aplasia o hipoplasia de
la hipfisis o de las adrenales.
Inmediatamente antes del parto aumentan los corticoides adrenales en el plasma maternal,
pero su efecto en el proceso del parto se considera imperceptible, en contraste con el
notable papel desempeado por la corteza adrenal del feto [1]. Es interesante conocer, que
Hipcrates 400 aos a.n.e., sugiri la importancia del feto en el inicio del parto.
FIGURA 5.1. Cambios en los niveles de progesterona, estradiol y PGF 2a en el plasma, al final de la
gestacin, en la cabra [4]
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
FIGURA 5.2 Cambios en los niveles de progesterona y de estradiol en el plasma, al final de la gestacin, en
la mujer [3].
El aumento de los corticoides fetales est asociado con la sntesis por los placentomas de
prostaglandina F2a y estrogenos, los que frenan la produccin de progesterona (Figura
5.3).
En la vaca los niveles de prostaglandina F2a en la vena tero-ovrica permanecen
relativamente constantes (< 0,5 ng/ml) hasta 5-7 das antes del parto, entonces se
incrementan de forma gradual alrededor de 1,5 ng/ml de 24-48 h antes del parto. La
concentracin de prostaglandina se incrementa bruscamente durante las ltimas 24-48 h de
preez a un mximo de 5,5-9 ng/ml durante el parto [6].
Los estrgenos aumentan durante la preez en los animales domsticos y alcanzan un pico
pocos das antes del parto. En la vaca los niveles de estradiol se incrementan ms o menos
en valor de 30 pg/ml sobre los ltimos 30 das de preez a alrededor de 300 PG/ml dos das
antes de producirse el parto. La concentracin disminuye a alrededor de 50 PG/ml un da
despus de este [7]. Estos niveles aumentados de estrgenos en la preez causan un
crecimiento del miometrio, sntesis de actomiosina y consecuentemente un incremento de
la capacidad contrctil del tero. Un descenso de progesterona con predominio absoluto o
relativo de estropenos, produce un aumento del potasio intracelular y del sodio
intersticial.
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
FIGURA 5.3 La sntesis de estrgenos y PGF2a por los placentomas frenan la produccin de progesterona
[5]
La actomiosina contenida en las mioclulas, bajo la accin del cido adeno-sintrifosfrico
(ATP) y en presencia del potasio y el magnesio, trae como consecuencia la aparicin de un
incremento de la excitabilidad muscular del miometrio. Un desequilibrio estrgenoprogesterona en el sentido de un predominio de los primeros es una de las causas
reconocidas hoy como desencadenantes del parto. Tambin los estrgenos junto a la
relaxina causan una relajacin del canal de parto, especialmente del crvix y la vagina [1].
Prximo al parto el tero es crticamente sensitivo a bajas concentraciones de oxitocina. En
muchas especies hay una sbita liberacin de oxitocina justamente antes del final de la
expulsin del feto. En la vaca se ha observado que la distensin de la vagina causa un
incremento en la liberacin de oxitocina, necesaria para la expulsin completa del feto
(figura 5.4). Lo mismo ha sido observado en la cabra. Sin embargo, en los animales
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
domsticos la oxitocina carece de importancia en el inicio del parto, con excepcin de la
yegua, especie en la que es capaz de inducirlo.
La intervencin del sistema nervioso central ha sido estudiada y se ha comprobado que la
denervacin uterina no puede impedir que se produzca el parto. No obstante, es probable
que un sistema nervioso central intacto mejore el proceso de este [1].
Se puede concluir que en los animales domsticos durante la gestacin, un alto nivel de
progesterona mediante la accin de la bomba de sodio mantiene sedado al tero. El primer
elemento de la cadena en el desencadenamiento del parto fetal, se sita en el hipotlamo
fetal, el que ante una situacin de estrs segrega el corticotropn RH. Este estimula la
secrecin de AGTH hipofisaria, que actuando sobre la adrenal fetal, da la seal de inicio
del parto, por medio de una descarga de cortisol.
El cortisol acta sobre los placentomas, los que incrementan los niveles plasmticos de
prostaglandina F2a y de estrgenos, que frenan la produccin de progesterona, por lo que
se establece un desequilibrio estrgeno-progesterona a favor del primero (Figura 5.5). Este
predominio de los estrgenos, incrementa la excitabilidad del miometrio, desencadenando
el parto. Por ltimo, al avanzar el feto dilata la vagina, lo que provoca una sbita liberacin
de oxitocina, que contribuye a la expulsin completa de este.
FIGURA 5.4 Concentraciones de oxitocina en dos vacas antes, durante y despus del parto [8]
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
FIGURA 5.5 Diagrama sumario que muestra los factores ms importantes envueltos en la regulacin del
parto [9]
Signos de la proximidad del parto
Los prdromos o manifestaciones que preceden al parto tienen gran importancia en la
preparacin de las condiciones para la posterior expulsin del feto y sirven, adems, para
pronosticar aproximadamente el momento en que se desarrollar el parto.
Dentro del desarrollo de la fase preparatoria del parto son notables las modificaciones
hormonales que se producen, las que inducen cambios qumicos en los tejidos de los
rganos genitales e influyen en la circulacin y la permeabilidad de los vasos sanguneos,
provocando la relajacin de los tejidos blandos en la regin pelviana.
Uno de los signos ms caractersticos es la relajacin de los ligamentos sacrociticos, que
se manifiesta en algunos animales por una ligera cada o hundimiento de los msculos que
cubren esta regin, La porcin de estos ligamentos que ms notablemente se relaja es la
parte situada entre las vrtebras coccgeas y las tuberosidades isquiticas, borde posterior
del ligamento, lo que provoca que la base de la cola (masto) aparezca ms elevada que lo
habitual.
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
Pocos das antes del parto es posible, aplicando una presin suave, desplazar el borde
posterior del ligamento aproximadamente 2,5 cm. La relajacin completa del borde caudal
de los ligamentos plvicos, por lo general, es acompaada del parto en un perodo de 12 h.
Su palpacin interna y externa es un mtodo seguro para pronosticar la inminencia del
parto.
El comienzo de la relajacin progresiva de los citados ligamentos coincide con el inicio del
ablandamiento y la relajacin del cuello uterino. Conjuntamente a este proceso se produce
tambin la tumefaccin, la edematizacin y la flacidez de la vulva, la que pierde
progresivamente el sistema plegable y puede alcanzar de 2-4 veces su volumen normal,
especialmente en la novilla.
Otro signo premonitor del parto lo constituye la salida por la hendidura vulvar de un moco
denso, viscoso y de color amarillento opaco, que con frecuencia forma cordones gruesos
(grosor de un lpiz o ms). Estos cordones penden de la vulva y alcanzan, en ocasiones,
una longitud de 30-40 cm. El moco procede de las secreciones que tienen lugar en el cuello
uterino y la dilucin del tapn mucoso de la gestacin, este proceso de eliminacin del
tapn mucoso se inicia en las novillas dos o, en ocasiones, tres semanas antes del parto y
ocurre como consecuencia de la dilatacin de la porcin vaginal del cuello uterino, es fcil
observar a la inspeccin vaginal, una dilatacin de 4-6 cm en el primer anillo, mientras el
resto del conducto permanece todava cerrado.
Las citadas secreciones son abundantes en la vaca de 6-15 das antes del parto, adquiriendo
aspecto de clara de huevo; quedan adheridas a los alrededores de la vulva y a la cola por
los movimientos de esta al tratar de eliminarlas. Los campesinos dicen entonces que la
vaca est limpiando para parir. El papel esencial de estas secreciones es el de lubricar el
canal del parto y reblandecer sus paredes para facilitar su distensin al pasar el feto.
Los cambios producidos en la glndula mamaria al final de la preez, son elementos de
valor en el pronstico de la proximidad del parto, esta glndula sufre una serie de
modificaciones anatmicas y fisiolgicas, siendo la ms evidente el incremento de
volumen, el que en las vacas se hace manifiesto 2-3 semanas antes del parto. En las
novillas se aprecia un desarrollo incipiente en el cuarto mes de la gestacin.
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
En las primparas y en vacas de alta produccin se producen edemas como consecuencia de
trastornos circulatorios que provocan la tumefaccin de la glndula y la acumulacin de
una considerable cantidad de lquido (plasma) en el tejido celular subcutneo,
especialmente en la regin umbilical donde algunas veces semeja una hernia. En las
novillas, en ocasiones, estos edemas son muy pronunciados (anormales) y pueden llegar
hasta la zona del esternn y extenderse a las extremidades borrndoles los contornos de las
articulaciones; excepcionalmente los edemas se presentan en la regin perineal.
Existe un signo que preludia la proximidad del parto en breves horas, que es la distensin
de los pezones, los que se tornan tensos. La secrecin de la ubre, serosa pegajosa, se torna
espesa justamente antes del inicio del parto y toma un color amarillento opaco (calostro), y
en casos de gran distensin, fluye al exterior.
Por ltimo, hay un signo que no se presenta en todas las hembras con la misma intensidad:
la disminucin de la temperatura corporal de 0,5-1 C, la que tiene lugar al disminuir la
tasa de progesterona por debajo de 3 mg en la circulacin perifrica materna. Una cada de
temperatura de alrededor de 0,5 C durante las ltimas 4P h antes del parto en las ovejas y
una por debajo de 39,2 C ha sido propuesta como un mtodo para seleccionar las que
deben parir en los prximos dos das. Este mtodo se ha mostrado 80 % efectivo.
Etapas del parto
El parto es un proceso continuo, pero ha sido dividido arbitrariamente en tres etapas
claramente definidas: de dilatacin cervical, de expulsin fetal y de expulsin de las
membranas fetales o alumbramiento.
Etapa de dilatacin cervical
La etapa de dilatacin cervical comienza con las contracciones de las fibras musculares
longitudinales y circulares del tero y termina con la dilatacin completa del cuello
uterino. Las contracciones uterinas llamadas pujos tienen la caracterstica de las fibras
musculares lisas, con difusin peristltica procedente de la parte craneal en direccin a la
caudal, son involuntarias, rtmicas y dolorosas, con descansos indoloros. Las contracciones
estn reguladas, de modo autnomo, por el sistema ganglionar uterino. Resultan dolorosas
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
porque la contraccin uterina comprime los nervios situados en la pared de la matriz y
secundariamente los del cuello uterino al dilatarse [10].
Las contracciones uterinas, por ser dolorosas, hacen que la palabra contraccin sea en este
caso sinnimo de dolor. Se originan por la cada del nivel de progesterona que bloqueaba
con su accin sedante las contracciones uterinas y el incremento de la excitabilidad
causada por altos niveles de estrgeno. La actomiosina contenida en las mioclulas bajo la
accin del cido adenosintrifosfrico (ATP) y en presencia de potasio y magnesio las
contraen, siendo el agente de la contraccin uterina. En la vaca la oxcitocina raramente es
liberada por la hipfisis en esta primera etapa y no es esencial para la induccin del parto.
Al inicio de la fase de dilatacin, las contracciones en la vaca, se repiten cada 15 min y
duran de 15-30 s, pero cuando el parto avanza son ms frecuentes (cinco contracciones
cada 15 min) y se prolongan de 80-100 s; tambin aumenta su intensidad. Entre las
contracciones, la presin uterina no desaparece, porque se mantiene el tono uterino
(capacidad de adaptacin muscular al contenido cavitario), esta presin, dbil al principio
del parto, aumenta segn avanza este y alcanza el mximo de intensidad en el perodo
expulsivo, con una media de 66 mm Hg. La presin ejercida por las contracciones uterinas
es equivalente a 33 mm Hg, que se aaden en cada contraccin a la que mantiene el tono
uterino.
Por palpacin rectal es posible reconocer dichas contracciones ya que durante la
contraccin el tero se endurece cada vez ms y no permite percibir las partes fetales.
Despus de la contraccin el tero se relaja haciendo posible de nuevo el reconocimiento
del feto.
En la etapa de dilatacin son escasos o estn ausentes los signos del parto, las vacas en el
pastoreo buscan generalmente un lugar aislado, le aumentan las secreciones vaginales
(limos) y presentan licuefaccin y expulsin del tapn mucoso cervical.
Usualmente no son aparentes los signos de intranquilidad y de molestia hasta que el cuello
del tero se ha dilatado lo suficiente para permitir que pase una mano [5], Un arqueamiento
ligero de la regin dorsal es aparente en este momento. En las vacas pueden no ser
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
evidentes los sntomas de dolor abdominal, especialmente en las plurparas. En las novillas
es usual observar sntomas de clico e intranquilidad.
Las vacas al sentir dolor en el vientre observan su abdomen, defecan y orinan con
frecuencia especialmente en los momentos en que los sacos fetales entran en la pelvis.
Durante las contracciones el pulso materno se acelera, el ritmo del corazn fetal se lentifica
y el tero sufre una ligera isquemia. La vaca se echa y se levanta con frecuencia, arquea el
lomo, y levanta la cola.
El cuello uterino es ms firme y duro que la pared del rgano, debido al mayor contenido
de tejido conectivo (colgeno); la dilatacin del cuello se debe ms a cambios en las
caractersticas fsicas del colgeno cervical (maduracin) que a un aumento de la presin
intrauterina, algo bien demostrado en especies como la ovina, la caprina y la bovina, las
que tienen un cuello rgido [11]. Su reblandecimiento e incrementada dilatabilidad son
regulados hormonalmente. En la cerda, la principal hormona implicada en su regulacin
parece ser la relaxina, la que es secretada por las clulas lutenicas de sus cuerpos lteos al
final de la gestacin y se almacenan en estos [12].
La relaxina se libera en la sangre aproximadamente dos das antes del parto, con picos de
14-22 h antes que este se produzca, esta actividad se pierde rpidamente despus del parto
[13].
En la vaca se conoce poco sobre la regulacin hormonal de la dilatacin del cuello uterino
durante el parto. La relaxina tambin ha sido aislada en los ltimos estadios de la preez,
de sus cuerpos lteos [11] y de las placentas, pero es mucho menos activa que en la cerda.
Los principales cambios del tejido conectivo del cuello en el momento del parto son: la
separacin de los paquetes de fibras colgenas, el incremento en los proteoglucanos
intracelulares y el contenido de agua, al parecer como resultado de la despolimerizacin
proteoltica del colgeno, ya que no hay aumento de colagenasa [14]. Tambin hay un
incremento de glucosamino-glucanos. Mediante los citados cambios queda abolido el
cierre cervical y el tono muscular, mantenidos durante la gestacin por el sistema nervioso
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
vegetativo. Se produce as una prdida de tono y con ella se llega a la relajacin del
conducto del parto, particularmente del cuello uterino [15].
La porcin vaginal del conducto cervical comienza a dilatarse en las vacas
aproximadamente una semana antes del parto, permitiendo la entrada de 2-4 dedos. En las
novillas el conducto cervical permanece cerrado hasta el da anterior al parto y al iniciarse
este, el conducto es influido por las contracciones de las fibras musculares del tero,
dilatndose el orificio externo hasta alcanzar de 3-6 in. De 2-4 h ms tarde es que
comienza la dilatacin del orificio interno, lo que permite que se proyecte en este el
alantocorion. De 6-12 h despus de haberse iniciado las contracciones el cuello tiene de 67 in de ancho, constituyendo con la vagina un canal continuo que se encuentra lleno con el
alantocorion distendido.
En la etapa de dilatacin, la actitud del feto tiene que modificarse, pasando de una actitud
de flexin mxima de la cabeza y las extremidades que mantena durante la gestacin para
adaptarse al menor espacio posible en el tero, al de extensin de la cabeza y de las
extremidades anteriores o posteriores, de acuerdo con la presentacin, lo que posibilita su
salida a travs del estrecho canal del parto.
El proceso de extensin de la cabeza y de las extremidades se debe, principalmente, a las
contracciones peristlticas del tero, que la orientan en su direccin, o sea, hacia la pelvis.
Adems, intervienen los movimientos activos del feto, consecutivos a un enriquecimiento
del C02 en la sangre fetal [10].
En dicha etapa la longitud del tero disminuye y el feto precedido por porciones del
alantocorion y del amnios, es empujado adentro del conducto cervical. Bajo el impulso de
la creciente presin endouterina, la bolsa alantoidea avanza por la vagina y sale al exterior
a travs de los labios vulvares (Figura 5.6).
Fisiolgicamente el alantocorion no puede penetrar muy adelante en el conducto cervical
por impedrselo sus relaciones placentarias alrededor del orificio uterino interno. Por lo
general la rotura de esta bolsa, llamada bolsa de las aguas, se produce entre los labios
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
vulvares y algunas veces dentro de la vagina, porque la presin externa y la tensin del
lquido que encierra es superior a la resistencia de las membranas que la constituyen.
La duracin de la etapa de dilatacin en la vaca es aproximadamente de 6 h y algo ms
larga en las novillas, aunque resulta difcil de precisar ese perodo por la imposibilidad de
determinar con exactitud el momento de su inicio.
La dilatacin que produce en la vagina la penetracin en ella de la bolsa alantoidea, unido
al cierre de la glotis, provocan contracciones reflejas de los msculos abdominales y del
diafragma, lo que marca el inicio de la segunda etapa del parto o etapa de expulsin.
Se debe tener presente que en la primera etapa, se dilata el canal cervical, pero no se
completa la dilatacin del canal del parto, puesto que el anillo himenial se dilata durante la
etapa de expulsin, por accin mecnica del feto.
FIGURA 5.6 Ha comenzado la fase de expulsin y la posicin echada de la vaca ha facilitado la salida al
exterior de la bolsa alantoidea
Etapa de expulsin letal
Esta etapa del parto se caracteriza por: ruptura de la bolsa alantoidea, entrada del feto en el
canal del parto -con lo que concluye su dilatacin-, contracciones abdominales (prensa
abdominal), ruptura de la bolsa amnitica y expulsin del feto por la vulva. Las
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
contracciones uterinas en esta etapa incrementan su frecuencia e intensidad y son
reforzadas por las contracciones abdominales, que proveen la fuerza adicional necesaria
para la expulsin del feto.
Las contracciones de la prensa abdominal solo se presentan cuando penetran partes fetales
en el canal del parto, las que comprimen puntos del cuello uterino y de la vagina.
Al comienzo de esta segunda etapa del parto, las contracciones se producen cada 3-5 min,
y se hacen ms frecuentes en la medida en que el proceso del parto progresa, hasta que se
ejercen cada 1 1/2-2 1/2 min.
Durante una de las primeras contracciones de la prensa abdominal el saco corioalantoideo
que se ha acercado a la vulva se rompe. Si el animal est parado, por lo general, estalla
cuando an est en la vagina. En los casos en que el animal se encuentra echado, la presin
intraabdominal fuerza al citado saco corioalantoideo a travs de la vulva, donde en una de
las contracciones se rompe.
El grado en que se proyecta hacia el exterior, depende hasta cierto punto del nmero de
cotiledones que se hayan separado de las carnculas [16]. Con la ruptura de la bolsa
alantoidea sale al exterior parte del lquido que contiene y disminuye la presin que la
bolsa ejerca en las paredes de la vagina y vulva, con lo que declina o cesa temporalmente
el esfuerzo abdominal.
En este pequeo receso, el saco amnitico ms resistente, es forzado a travs del cuello
uterino y aparece en la vulva como una estructura blanca, opaca, avascular bolsa de las
patas. (Figura 5.7). Despus de esto la vaca se echa y se levanta no manteniendo por
mucho tiempo ninguna de estas posiciones. La salida del ternero puede ser completa dentro
de las 2 h siguientes al momento en que el saco amnitico aparece en la vulva [16].
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
FIGURA 2.7 La bolsa alantoidea se ha roto y aparece la amnitica la que se pueden observar las patas del
ternero.
Tras la pausa que sigue a la ruptura de la bolsa corioalantoidea se produce un repentino
incremento de las contracciones abdominales que se superponen a cada contraccin
uterina. Estas contracciones continan y las extremidades anteriores del feto, o en algunas
ocasiones, las posteriores, aparecen en la vulva, contenidas en la bolsa amnitica. Cuando
las extremidades pasan a travs de la vulva, la bolsa amnitica ha llegado a los lmites de
sus posibilidades de exteriorizarse y, generalmente, se rompe (Figuras 5.8 y 5.9), ante la
presin de las extremidades fetales, dejando escapar parte del lquido que contiene, el que
tiene propiedades lubricantes. Despus de la ruptura del amnios, el animal trata de lamer
tanto lquido como le sea posible. El intervalo promedio entre la ruptura de la bolsa
corioalantoidea y la amnitica es alrededor de 1 h [17].
Luego de otro breve descanso que sigue a la ruptura de la bolsa amnitica el esfuerzo
abdominal se restablece de nuevo y aumenta progresivamente en frecuencia hasta llegar en
muchos animales a ser constantes en los finales del nacimiento.
Al acercarse el feto a la vulva la vaca se echa durante largo tiempo. Entre los perodos de
esfuerzo, el animal descansa sobre el esternn con los miembros flexionados y rumiando o
comiendo espordicamente, o con la cabeza hacia el flanco [13]. Cuando inicia cada
esfuerzo abdominal se pone de lado, extendiendo las patas, y en el punto mximo de la
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
contraccin eleva la cabeza y las extremidades posteriores, dirigindolas hacia delante,
modificando en esta maniobra la posicin de la circunferencia pelviana o estrecho anterior
de la pelvis, con el fin de que la conjugada verdadera tome una posicin favorable que
permita el avance del feto (ver Figura 5.7). Si el animal se encuentra de pie, inclina
igualmente su pelvis en cada contraccin, con el mismo fin. Esto lo logra encorvndose,
con lo que baja la grupa.
FIGURA 5.8 La cabeza y las extremidades anteriores han salido al exterior. La presentacin es ceflica, la
posicin dorso sacra y la actitud de extensin (euttica)
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
FIGURA 5.9 Una extremidad posterior ha salido al exterior. La bolsa amnitica se ha oto y vierte lquido
que lubrica el canal del parto.
El mayor retraso en la expulsin del feto se produce al llegar su cabeza a la vulva. En este
punto, el progreso de expulsin por las contracciones uterinas es muy poco, pues el feto se
mueve continuamente hacia la vagina entre los perodos de contraccin. Este hecho se
acenta cuando la madre est en pie y con frecuencia las patas vuelven a la vagina [16].
Para forzar la cabeza a travs de la vulva, generalmente se requiere un mayor esfuerzo de
expulsin.
Cuando la cabeza se introduce entre los labios vulvares el esfuerzo tiene que ser
considerable para vencer ese, el mayor obstculo, pues el occipucio fetal queda detenido
por la comisura superior de la vulva y, adems, en ese momento est entrando el trax fetal
en el canal del parto. Es entonces cuando se producen las mayores y ms fuertes
contracciones abdominales en el proceso del parto [18] debidas en buena medida a la
continua descarga de oxitocina que provoca la dilatacin de la vagina.
A menudo, despus que la cabeza pasa la vulva, la madre descansa unos minutos antes de
comenzar de nuevo fuertes contracciones abdominales, para forzar el paso del tronco del
feto a travs del canal del parto y la vulva. El dimetro externo-dorsal del feto excede en
cerca de 7 cm al sacropubiano de la madre, por lo que las apfisis espinosas de las
vrtebras torcicas tienen que inclinarse hacia atrs y abajo, unas sobre otras, al mismo
tiempo que el esternn retrocede y arrastra en el mismo sentido la porcin distal de las
costillas que se insertan en el mismo [10].
El paso por la pelvis del cinturn pelviano del feto, encuentra una ligera dificultad, ya que
la distancia entre sus trocnteres mayores supera en 2 cm aproximadamente al dimetro
bisiliaco superior de la madre, pero esta dificultad es resuelta fcilmente al rotar el feto en
su salida unos 45 (Figura 5.10), tanto en los casos en que la madre est de pie, como
cuando est echada, con lo que los trocnteres pasan por uno de los dimetros oblicuos de
la pelvis, los que en la vaca son mayores que el transverso, por ser su pelvis dolicoplvica
(Figuras 5.11 y 5.12).
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
FIGURA 5.10 El tronco ha salido y el ternero ha rotado 45
FIGURA 5.11 La rotacin del ternero permite que su dimetro bisiliaco atraviese el estrecho anterior de la
pelvis por su dimetro oblicuo el mayor
FIGURA 5.12 La madre al pararse facilita la salida completa del ternero
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
Cuando se interviene en el parto, aplicando traccin forzada, es preciso tener en cuenta la
necesidad de rotar el feto despus de la salida del trax, ya que si no se toma esa medida, el
cinturn pelviano del feto se encaja en la pelvis e impide su salida, Esto, en ocasiones, es
causa de distocia provocada, en partos atendidos por legos.
El cinturn pelviano, en partos eutcicos no atendidos, usualmente atraviesa el canal del
parto rpidamente mediante la rotacin espontnea del feto, pero a veces las extremidades
posteriores permanecen en la vagina y el cordn umbilical se mantiene intacto, hasta que
se rompe por un movimiento de la madre o del recin nacido.
La ruptura del cordn umbilical con sus vasos sanguneos y el uraco se origina a una altura
de 10-20 cm de la pared abdominal. Tras su rotura generalmente no hay sangramiento, al
producirse el colapso de las venas y la retraccin de los muones arteriales hacia la
cavidad abdominal, debido a la contraccin de las fibras elsticas, muy abundantes en la
pared vascular y a causa de la unin laxa de los vasos, los tabiques del cordn y el anillo
umbilical. Es usual que solo se produzca sangramiento en los casos en que el cordn se
rompa en su lmite con la piel.
La duracin de la etapa de expulsin, en las multparas, es de 0,5-2 h y en las nulparas
alcanza de 3-4 h. La expulsin del feto, cuando el animal no es molestado y solo recibe
ayuda en el estadio final del parto, se produce en la generalidad de los casos en posicin
echada de la madre (ver Figuras 5.6, 5.12, 5.13 y 5.15)
En algunos casos, en la etapa de expulsin sale primero la bolsa amnitica, y la alantoidea
aparece todava intacta durante el alumbramiento, en la hendidura vulvar, de este modo se
cree a veces errneamente, en la existencia de un segundo feto.
Etapa de expulsin de las membranas fetales o alumbramiento
El alumbramiento es el proceso fisiolgico de desprendimiento y expulsin de las
membranas placentarias o secundinas. En el ganado bovino este proceso tiene una duracin
aproximada de 4-8 h, considerndose fisiolgica una demora de hasta 12 h, despus de la
expulsin del feto.
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
Desde los ltimos meses de la gestacin, la placenta comienza a sufrir cambios
preparatorios que facilitan su posterior desprendimiento, estos tienen lugar en su epitelio y
tejido conectivo. Un importante cambio es la disminucin de las clulas binucleadas
gigantes, en el trofoblasto, al final de la gestacin, la que ha sido observada en la oveja y
en la vaca [19] [20] y una disminucin de su nmero, observada 1 h despus del parto, en
los animales que expulsan la placenta antes de las 12 h del parto [21].
En la etapa de dilatacin, y particularmente, durante las contracciones del tero hay un
cambio constante de presin uterina que comienza a alternar condiciones anmicas e
hipermicas y cambios temporales en la superficie de las vellosidades corinicas. Como
resultado comienza a afectarse la unin del epitelio de las vellosidades con las criptas
uterinas [22]. En el perodo de expulsin es evidente, en la periferia de las carnculas, el
primer signo del proceso de desprendimiento mecnico, probablemente como consecuencia
de la presin que estas sufren contra el feto en cada contraccin uterina [22].
Un factor esencial, despus de la expulsin del feto y la rotura del cordn umbilical, es la
anemia de las vellosidades coriales, como resultado de la prdida de circulacin sangunea
en sus capilares, los que se colapsan, reduciendo con ello de manera considerable el
volumen de las vellosidades, hecho que favorece su desprendimiento de las criptas de las
carnculas.
Las contracciones uterinas que suceden a la expulsin del feto (entuertos) completan el
proceso de desprendimiento de la placenta y facilitan su expulsin.
Nomenclatura obsttrica
Ha sido necesario establecer una nomenclatura para describir las distintas presentaciones,
posiciones y actitudes que puede asumir el feto cuando penetra en el canal del parto, la que
es utilizada en la descripcin del parto, tanto el eutcico o normal, como el distcico o
difcil. El uso de esta nomenclatura resulta indispensable para la comunicacin entre
profesionales.
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
Presentacin
Expresa la relacin que existe entre el eje longitudinal del feto o eje cerebros-pinal y el
canal maternal del parto. Las presentaciones normales o eutcicas son las longitudinales,
las que pueden ser anterior o ceflica, que es la ms frecuente y ms favorable para el
parto, y la posterior o caudal, segn la porcin del feto que se presenta en el canal del
parto. Las presentaciones distcicas son las transversales, las que pueden ser dorsal o
ventral y la vertical, que es muy rara y solamente la vertical oblicua posicin de perro
sentado de la especie equina merece consideracin [23]. En la vaca esta presentacin es
extremadamente rara.
Posicin
Indica la relacin del dorso del feto con un punto del cinturn plvico de la madre [10]
[18]. La posicin eutcica es la dorsosacra y las distcicas son la dorsopubiana y las
dorsoiliacas derecha e izquierda. Algunos autores establecen la relacin del dorso fetal con
el canal del parto y le llaman posiciones dorsodorsal, dorsoventral y laterales derecha e
izquierda [23], otros [24] denominan posicin superior a la dorsosacra y posicin inferior a
la dorsopubiana.
Actitud
Se refiere a la disposicin de los apndices mviles del feto, los que se pueden encontrar
en extensin que es la actitud normal o eutcica para el parto, o en flexin de alguno de
estos apndices, que constituyen actitudes distcicas. Durante la gestacin todos los
apndices mviles del feto se encuentran en actitud de flexin mxima, para adaptarse al
menor espacio posible en el claustro materno.
Durante el parto distcico, las actitudes de flexin de la cabeza, pueden ser la flexin
ventral o las laterales derecha o izquierda. La flexin dorsal de la cabeza solo se presenta
en especies de cuello largo. Las actitudes de flexin de las extremidades en la presentacin
anterior, pueden ser del carpo o del encuentro y en la presentacin posterior del tarso o
corvejn, o de cadera. Todas ellas pueden ser unilaterales, o bilaterales si la flexin
comprende ambas extremidades. La actitud de cadera en flexin bilateral es conocida
como presentacin de nalgas.
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
Transcurso del parto
Es conveniente permitir que el parto transcurra libremente, mientras que no se interrumpa
o altere alguna de sus etapas. Se debe tener en cuenta que el canal del parto no se encuentra
previamente formado, sino que se va conformando por la presin ejercida por las
extremidades y la cabeza fetal en su trnsito hacia el exterior y solo queda constituido en
su totalidad cuando estas acaban de salir por la vulva, y el cinturn escapular y el tronco lo
estn atravesando.
El parto fisiolgico es un proceso que se desarrolla lentamente, sin que el feto reciba dao
alguno, pero en muchas ocasiones, personal inexperto aplica traccin innecesariamente,
antes de que el canal blando est formado, y cuando la traccin es intensa, el feto sufre y su
vitalidad se afecta.
Cuando el parto deja de transcurrir de modo normal, s se impone un rpido diagnstico,
para conocer cul es la situacin y brindarle ayuda si esta es necesaria. Determinar cundo
se ha interrumpido el curso normal del parto, lo permite la experiencia acumulada, debido
a la observacin y a la asistencia a un crecido nmero de partos. Si la experiencia falta, o
resulta insuficiente, es til apoyarse en la Gua para la intervencin en el parto ofrecida por
Schuijt y Ball [25]. Segn esta gua, se debe realizar un examen diagnstico en las
situaciones siguientes:
1. Cuando han transcurrido ms de 6 h de comenzada la fase de dilatacin y no se han
presentado contracciones de la prensa abdominal, es decir, no ha comenzado la segunda
fase, o fase de expulsin fetal.
2. Cuando la hembra lleva 2 o 3 h en la fase expulsiva y no se nota, o se nota muy poca
progresin en el parto.
3. Cuando han transcurrido 2 h despus de observarse la bolsa amnitica y no se ha
producido el parto.
4. Tambin es necesario examinar a la hembra cuando hace ms de 1 h que se rompi la
bolsa alantoidea y no se ha presentado en la vulva la bolsa amnitica.
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
Examen diagnstico de la parturienta
El diagnstico inicial que se realiza a la hembra que se presume presenta alguna dificultad
en el parto, se debe practicar con estrictas medidas de higiene, pues a veces, solo es
necesaria esta intervencin en el parto.
Nunca se debe olvidar que la asistencia profesional al parto, tiene que caracterizarse por la
observacin de una correcta higiene. La vulva y el resto de la regin perineal y, en especial,
el maslo de la cola se deben lavar cuidadosamente con agua y jabn, antes de proceder al
examen. El obstetra se debe lavar las manos y ponerse un guante para disminuir la
contaminacin y protegerse de las enfermedades infecciosas. Desdichadamente los guantes
no se pueden utilizar para manipulaciones prolongadas porque se rompen con facilidad.
El examen debe realizarse con la vaca parada, ya que presenta muchas dificultades cuando
se practica con el animal en el suelo, por el aumento de la presin abdominal. Si el animal
est echado debe ser instado a que se pare, lo que se logra en un buen porcentaje de
animales. Algunos estn exhaustos o no desean pararse y requieren mtodos ms vigorosos
como pincharlos con un objeto duro y puntiagudo, doblarles la cola (sin partrsela), darle
con la rodilla en el costillar, aplicarle una garrocha elctrica o si no una inyeccin
endovenosa de algn estimulante del sistema nervioso central, como la anfetamina, la
coramina o la piribenzamina, si los mtodos anteriores no dan resultado.
En ocasiones, despus de aplicarles anestesia epidural, animales que se mantenan en el
suelo se paran, al cesar el dolor. Si no se logra que el animal se pare es conveniente
elevarle el tren posterior por medio de una puerta o de un tabln grueso calzado con
bloques, Esto se logra tambin colocndole pacas de heno.
El animal se debe mantener sujeto en forma apropiada para impedir que dae al operador.
Se debe disponer de agua corriente o de varios cubos de agua y de una mesa que se cubre
con un pao estril para colocar el instrumental.
Se comenzar observando la vulva, su edematizacin, dilatacin, y si tiene aspecto de
haberse hundido. A continuacin se debe examinar con cuidado el canal blando del parto,
prestando especial atencin a la dilatacin cervical y la de las estructuras de la regin
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
vestibular anillo himenial, constituido por el constrictor posterior de la vulva, msculo de
fibras estriadas reforzado por haces musculares del elevador del ano- Se determinar
cualquier posible anomala o deformacin del estrecho anterior de la pelvis, as como se
localizar el feto. Si este se encuentra en la cavidad pelviana, se examina para determinar
su presentacin, posicin y actitud, as como cualquier anomala teratolgica. En algunas
ocasiones se pueden confrontar dificultades para distinguir si una extremidad es anterior o
posterior. Si la cara plantar est hacia abajo es muy probable que sea anterior y en caso
contrario posterior, pero el diagnstico de certeza requiere que se observe la direccin de la
flexin de sus articulaciones. Si la articulacin por encima de la articulacin del menudillo
se flexiona en su misma direccin es anterior y si se flexiona en sentido contrario es
posterior.
Se debe determinar si el feto est vivo o muerto, ya que su estado influye el pronstico y la
decisin sobre el proceder obsttrico. Cuando el feto est vivo, si se toma una extremidad y
se le aprieta o se le empuja, esta se mover. Si abrigamos alguna duda se pueden examinar
los reflejos, en la presentacin anterior se comprobarn el reflejo ungular, el ocular o el de
succin. Algunas veces la introduccin del dedo en la boca provoca movimientos de la
lengua y de la mandbula.
En la presentacin posterior se pueden comprobar el reflejo ungular o el anal. En ausencia
de estos reflejos se puede palpar el trax para percibir los latidos cardacos o el cordn
umbilical, para sentir el pulso, antes de llegar a una decisin final sobre el estado vital del
feto.
Si el feto est muerto, es necesario conocer si presenta descomposicin y su grado, lo que
se sabe por la presencia e intensidad del enfisema, por la cada del pelo y por el olor, que
puede ser ptrido.
Durante este examen tambin se establece la relacin feto-plvica, con el objetivo de
conocer si el feto puede alcanzar el exterior por la va vaginal. Para ello se pueden utilizar
mtodos pelvimtricos, como la frmula establecida por Hindson [26].
Cuando el obstetra tiene amplia experiencia, el grueso de la articulacin del menudillo le
es suficiente en muchos casos para establecer la mencionada relacin.
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
La Relacin de Traccin (RT) se refiere a la medida de la distancia nter-isquitica dividido
con la del menudillo (DM) del ternero, con el que tiene una correlacin altamente
significativa. Para tomar la primera medida us una regla de 20 cm y para la segunda un
calibrador.
Al dividir la separacin entre las tuberosidades isquiticas, entre el dimetro de la
articulacin del menudillo, obtuvo un cociente que denomin la relacin de traccin:
STI
RT
DM
Despus de establecer la validez de estas medidas bsicas correlacion RT con el resultado
de casos de distocia que tuvieron cras muertas o en las que haba posibilidad de presentar
una desproporcin fetoplvica, llegando a las siguientes conclusiones:
1. Cuando RT es mayor que 2,5 no hay desproporcin fetoplvica.
2. Cuando RT se encuentra entre 2,3 y 2,5 puede haber desproporcin fetoplvica, pero se
obtiene resultado con la traccin.
3. Cuando RT est presente entre 2,1 y 2,3 la traccin puede tener dificultades y se debe
considerar la cesrea como solucin.
4. Cuando RT es menor que 2,1 es poco probable obtener un ternero vivo por traccin,
por lo que la cesrea debe ser el mtodo de eleccin.
En Los casos que manifiesten dificultades adicionales, como es el de las novillas, las que
se pueden presentar fallos en la relajacin de los tejidos o inmadurez, los casos de
presentacin posterior y el de las razas que muestran hipertrofia muscular, se requiere la
aplicacin de factores de correccin a RT, estos son:
Para la separacin entre las tuberosidades isquiticas 577;
N - novillas, factor de correccin 0,95
Para el dimetro del menudillo DM:
PP - presentacin posterior, factor de correccin 1,05
HM - hipertrofia muscular, factor de correccin 1,05
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
Para estos casos se tiene, por consiguiente, la frmula:
RT
STI N
1
.
.
DM PP HM
Nacimiento del feto
El feto al nacer, en su trnsito forzado al exterior, tiene que vencer dificultades que en
determinados momentos son extremadamente serias, e inmediatamente debe afrontar
enormes cambios. Del medio muy favorable en que se desarroll dotado de un eficiente
sistema de proteccin y una temperatura constante, con muy ligeras fluctuaciones pasa a
uno hostil, donde recibe agresiones de todo tipo. Su circulacin placentaria, en segundos
pasa a pulmonar, y lo ms neurlgico, debe iniciar rpidamente la respiracin pulmonar, su
pena de no alcanzar la existencia.
Inicio de la respiracin pulmonar
Sin dudas el comienzo de la respiracin pulmonar y su establecimiento como respiracin
definitiva es el momento ms crtico del nacimiento del feto. Por ello cuando se asiste al
parto, se debe tratar de minimizar las dificultades desde su inicio y favorecer su
establecimiento.
El primer paso es liberar los ollares de la obstruccin que pueden causar restos de
membranas fetales. El mucus y los fluidos fetales deben ser extrados de los ollares y la
boca, por presin externa de los pulgares sobre los cornetes nasales, con el resto de los
dedos en el espacio intermandibular, deslizndose desde el nivel de los ojos en direccin al
morro [27],
Si la respiracin no ha comenzado se estimula su inicio, lo que se puede lograr con
frecuencia, vertindole agua fra sobre la cabeza, frotndote la piel enrgicamente con un
pao o hacindole cosquilla en la mucosa nasal con una pajita para provocarle un
estornudo. Tambin se puede utilizar un analptico respiratorio como la niquetamida a una
dosis de 0,1 g. La administracin de estos productos hace que el ternero realice varias
inspiraciones profundas. Como cada inspiracin es un estmulo para la prxima, estos
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
agentes pueden tener un efecto favorable. Estos estimulantes tienen un efecto de solo 5-10
min [27].
Despus de un parto que ha requerido asistencia, es posible que sea necesario extraer un
exceso de moco de la traquea y los bronquios. Esto se puede lograr mediante succin
practicada con un extractor, o suspendiendo el ternero por los miembros posteriores, con la
cabeza en el aire, durante 10-30 s. Se puede aplicar una ligera presin con las manos a lo
largo del trax desde las costillas hasta el cuello, o se puede balancear en crculo, para que
la fuerza centrfuga desarrollada ayude a evacuar el moco. Un balanceo severo o
prolongado puede tambin evacuar el contenido estomacal del ternero, el que podra ser
aspirado al suspender dicho tratamiento [18]. Se debe tener presente que estos tratamientos
no actan sobre el pulmn no desplegado del recin nacido.
RESPIRACIN ARTIFICIAL
Para aplicar la respiracin artificial, se requiere limpiar previamente la boca y los ollares,
de moco, y mantener la boca del ternero abierta con la lengua extendida para que el aire
pase libremente.
Se puede aplicar la tcnica descrita por Maarten Drost [27] segn la cual el ternero se
coloca en decbito lateral y el operador se sita detrs de este. Con una mano agarra el
hmero y la otra la coloca por debajo de la ltima costilla, entonces levanta la extremidad
y junto con ella la pared costal, hasta que el ternero se separa del suelo. Esto expande el
trax y da al pulmn oportunidad de expandirse. Esta expansin es ligera, porque los
pulmones estn atelectsicos; a continuacin se comprime firmemente la pared torcica
con las palmas de las manos. Estos movimientos se repiten aproximadamente uno cada 5 s
y se trata que el mayor esfuerzo se realice en la inspiracin.
Generalmente no se obtiene sonido expiratorio hasta despus de muchos movimientos de
resucitacin. Cuando el pulmn comienza a expandirse, es aspirado muy poco aire. Al
presentarse movimientos respiratorios expontneos deben ser ayudados, manteniendo la
ritmicidad de la respiracin artificial, la que se puede mantener por 15 min.
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
Otra tcnica para aplicar respiracin artificial ha sido expuesta por Sloss y Dufty [16], en
ella el ternero se coloca en decbito dorsal, con su cabeza y cuello dirigidos hacia abajo.
Primero se empujan los miembros anteriores en direccin medial para comprimir el trax,
y luego se separan para que se expanda. Este procedimiento debe continuarse de 5-10 min
y es importante mantener el ritmo de un movimiento cada 5 s. aproximadamente.
Despus que la frecuencia respiratoria ha alcanzado un nivel adecuado y el ternero ha sido
secado frotndolo, se debe situar sobre su trax con las extremidades anteriores extendidas,
separadas hacia afuera, y las posteriores en posicin de perro sentado, extendidas a lo
largo del cuerpo. Esta posicin es muy favorable para el ternero en esas circunstancias, ya
que facilita la expansin del trax. Es conveniente calzarlo por los lados con un mazo de
paja, para mantenerlo en la posicin en que se ha colocado, decbito esternal [27],
Estado cido-bsico del ternero recin nacido
Como consecuencia del proceso de su incorporacin al nuevo medio de vida, el ternero
recin nacido, aunque provenga de un parto normal, presenta una acidosis fisiolgica [28],
la que evoluciona de manera espontnea hacia el equilibrio, en las primeras 24 h de vida.
La expulsin prolongada o retardada del feto afecta los cambios gaseosos entre la
circulacin fetal y maternal. El oxgeno fijado y el dixido de carbono liberado son
insuficientes e inducen una acidosis respiratoria que se puedo convertir en metablica,
como resultado de la acumulacin de productos cidos del metabolismo, originados por
conversin anaerbica de las grasas [27].
Sloss y Dufty, reprodujeron este cuadro en forma experimental mediante el pinzado del
cordn umbilical, lo que fue seguido de la expulsin de terneros dbiles [16].
SUFRIMIENTO FETAL DURANTE EL PARTO
Despus de un parto difcil algunos temeros aparecen deprimidos con dificultad para
respirar. Una acidosis seria induce depresin central y vasoconstriccin que provoca una
rpida disminucin de la circulacin pulmonar [27]. Pueden presentar disnea o apnea,
flacidez muscular, pulso lento y reflejos disminuidos o ausentes, proceso este que
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
comienza por el ungular de las patas y contina por el ungular de las manos, el de succin
y termina por el corneal.
Otros terneros comienzan a respirar, pero son incapaces de levantar la cabeza y de moverse
para levantarse y mamar. Estn embotados y deprimidos, presentan hipotermia y pueden
manifestar tremor. Los reflejos de succin y
de deglucin dbiles o ausentes les
imposibilitan mamar. Su estado les hace fcil presa de trastornos respiratorios o
gastrointestinales. Estos terneros si no son adecuadamente tratados pierden peso con
rapidez y mueren de des-hidratacin e inanicin [6].
Una acidosis exactamente respiratoria mejora muy rpido con una apropiada ventilacin.
La acidosis metablica requiere una aplicacin intravenosa lenta de 40 o 50 mL de una
solucin estril de bicarbonato de sodio al 8 %. La dosis se repite si es necesario, a las 4 o
6 h.
La alimentacin forzada de terneros con reflejo de succin muy dbil, puede ocasionarles
una bronco-aspiracin de calostro. Adems, si el ternero est hipotrmico, su capacidad
para digerir y absorber los nutrientes est marcadamente reducida, por lo que es necesario
proveerles una fuente rpida de energa, que puede consistir en 500 mL de una solucin de
dextrosa al 5 % por va intravenosa.
ndice clnico del recin nacido
Olga Torres [29] elabor un ndice clnico muy til para evaluar el grado de sufrimiento
fetal, el cual consiste en la valoracin de los signos siguientes:
1. Levantar la cabeza (menos de un minuto).
2. Reflejo de succin (presencia, presin y continuidad).
3. Respuesta a los estmulos (pellizco en la grupa, agua en el odo y doblar la oreja
presionndola en el tronco).
4. Inters por el entorno (por observacin o con una palmada cerca).
5. Intento de pararse (varios intentos en menos de 20 minutos).
Puntuacin de cada signo:
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
Bien......... 2
Regular ....1
Mal..........O
Interpretacin:
Deprimido (0-5 puntos)............... acidosis grave o moderada.
Vitalidad afectada (6-7 puntos) ..acidosis leve.
Buena vitalidad (8-10 puntos) .....acidosis fisiolgica.
Ruptura del cordn umbilical
El cordn umbilical del ternero, el del cordero, el del cabrito y el del cerdito, por lo
general, se rompen cuando el feto pasa a travs del canal del parto. En el cerdito, por ser el
cordn relativamente largo, con alguna frecuencia se rompe por sus propios movimientos.
El cordn del potro, por ser ms largo y ms fuerte, comnmente se rompe por
movimientos de la madre para tratar de incorporarse despus del parto, lo que da
oportunidad a que una buena parte de la sangre de la placenta fetal pase a la cra. En los
carnvoros el cordn usualmente es cortado por la madre con sus dientes, dada su mayor
resistencia.
El desgarro natural, al igual que la mordedura cohben eficazmente la hemorragia
umbilical. Al romperse el cordn, los muones arteriales se retraen hacia la cavidad
abdominal, debido a la contraccin de las fibras elsticas que existen en abundancia en la
pared vascular, la vena umbilical se colapsa y el uraco se contrae.
Si el cordn umbilical no se ha roto por si mismo, no se debe cortar, sino romperlo por
traccin opuesta de ambas manos, cuidando que la ruptura no se produzca a nivel del
ombligo cutneo.
Tratamiento del cordn umbilical
Cuando el parto de la vaca se produce en un pasto limpio, no se requiere la desinfeccin
del ombligo [18]. Tampoco necesita tratamiento especial el ombligo de las especies
menores [24].
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
El mun umbilical puede ser desinfectado empapndolo con tintura de yodo, con una
solucin de tanino al 5 %. con una solucin de cido saliclico al 5% en alcohol al 70 %, o
con algn otro antisptico similar. Despus se comprime de arriba a abajo con un algodn
mojado en la solucin antisptica, para que la vena umbilical expulse el resto de sangre que
le pueda quedar.
Para que una cura umbilical con antispticos sea efectiva, se, requiere que sea
cuidadosamente aplicada, de 1-3 veces diarias, durante 2 o 3 das [7]. La cura debe ser a
cielo abierto y los restos del cordn se secan y caen, en 1 o 2 semanas.
Si por haberse roto el cordn umbilical muy prximo al ombligo cutneo persiste el
sangramiento y requiere una ligadura, se debe aplicar antibiticos de 3-7 das y remover la
ligadura a las 24 o 48 h.
En el potro a menudo es aconsejable administrar una dosis profilctica de antitoxina
tetnica. En potros de mucho valor se puede administrar antibiticos como penicilina,
estreptomicina o tetraciclina durante varios das para prevenir una infeccin del ombligo o
una septicemia.
Suministro de calostro
El ternero al nacer requiere se le provea de inmediato una inmunidad pasiva, para lo que es
esencial la ingestin de calostro, ya que este al igual que los potros, los corderos, los
cabritos y los cerditos, solo reciben inmunidad de la madre a travs de esa va. Los terneros
que reciben calostro tienen ms neutrfilos y estos tienen mayor actividad fagocitaria que
los privados de calostro. Adems, el calostro es rico en vitaminas A, D y E, que estn
presentes en niveles muy bajos en el ternero recin nacido [30].
El ternero debe consumir al menos el 6 % de su peso en calostro, en dos tomas, una
durante las dos primeras horas que siguen a su nacimiento y la otra 12 h ms tarde. Bajo
condiciones altamente insalubres o lugares infectados con grmenes virulentos, es
menester que reciban el calostro dentro de los primeros 15-30 min que siguen a su
nacimiento. Si es necesario mediante una sonda esofgica [18].
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
Las inmunoglobulinas circulantes protegen contra la septicemia, pero no siempre
previenen la diarrea, porque presumiblemente no alcanzan el lumen intestinal en
cantidades suficientes. No obstante, altos niveles en el suero de IgG e IgA, reducen la
severidad de la diarrea por prevenir un derrame masivo de fluidos y electrlitos al lumen
intestinal. Inversamente, altos contenidos de inmunoglobulinas en el intestino protegen
contra la diarrea, pero no contra la septicemia [31].
La urgencia requerida en el suministro del calostro, adems de por la necesidad de
conferirle una defensa inmediata al recin nacido, est dada, porque la nica posibilidad de
absorber las grandes molculas de inmunoglobulinas calostrales reside en ciertas clulas
epiteliales del intestino delgado (especialmente el yeyuno e leon), que desafortunadamente
tienen una corta vida, siendo sustituidas rpidamente por clulas maduras que no permiten
esa absorcin [1], y porque la riqueza de inmunoglobulinas del calostro solo est presente
el primer da, quedando al segundo da significativamente reducida (Tabla 5.1).
TABLA 5.1 COMPOSICIN APROXIMADA DE CALOSTRO Y LECHE NORMAL
DE VACAS HOLSTEIN [27]
Constituyente
Grasa
Primer calostro (%)
6
Calostro del 2do. y 3er. das
3.5
Leche (%)
3.50
Slidos no grasos
22.3
12.5
8.80
Protena
18.8
7.5
3.25
Inmunoglobulina
13.1
1.0
0.09
Seis horas despus del primer ordeo se ha perdido el 33 % de las inmunoglobulinas, por
dilucin, y despus de 24 h se pierden ms del 66 %.
La concentracin de inmunoglobulinas en la glndula mamaria comienza cinco semanas
antes del parto y alcanza mximos niveles aproximadamente tres semanas ms tarde [32],
El preordeo a que, en ocasiones, son sometidas las vacas con edemas severos de la ubre,
puede reducir las inmunoglobulinas disponibles en el calostro, ya que este solo es
secretado una vez en cada gestacin.
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
Las inmunoglubulinas contenidas en el calostro pueden ser cualificadas con un
calostrmetro, hidrmetro que mide la gravedad especfica del calostro. Una gravedad
especfica superior a 1,046 indica un contenido adecuado de globulinas.
Atencin a la vaca recin parida
Cuando se atiende un parto, aunque la asistencia va dirigida a extraer al feto y obtener una
cra viva y saludable, no se puede olvidar nunca que la madre generalmente es ms valiosa,
y preservar su vida, salud y fertilidad posterior, debe ser el objetivo econmico primario.
Despus de estar seguro que la respiracin del recin nacido ha quedado establecida, se
debe explorar el tero para descartar la presencia de otro feto. Afecta mucho al crdito del
profesional, retirarse despus de asistir un parto y darlo como concluido, y que la madre
posteriormente presente otro feto. An es mucho ms grave la situacin, si como ocurre
con alguna frecuencia, el segundo feto presenta una actitud distcica.
Previamente se debe lavar la vulva y la regin perineal con agua y jabn, as como el brazo
del operador, el que debe ser lubricado.
Cuando se explora el tero en busca de un segundo feto, se debe aprovechar para examinar
el canal del parto y detectar posibles lesiones o hemorragias. Traumas de los tejidos
blandos del tracto genital o de los huesos de la pelvis pueden provocar hemorragias fatales,
infecciones o incapacidad motora.
Traumas del canal del parto
Las heridas ligeras de la vulva, la vagina y el cuello frecuentes cuando se practica la
extraccin forzada, no requieren sutura, pero muchas veces se infectan, especialmente si
los tejidos estn traumatizados. Las heridas de la vagina por lo general se producen en las
paredes laterales, a nivel del anillo himenial, especialmente en las novillas gordas. A travs
de ellas en estas novillas se puede prolapsar grasa perivaginal, cuando se practica la
extraccin forzada de fetos grandes, sin que exista la dilatacin completa del esfnter
vestbulo vaginal.
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
La grasa prolapsada puede ser cortada con una tijera, lo que solo provoca un ligero
sangramiento. La herida de la vagina no necesita sutura [17]. El citado prolapso de grasa
perivaginal requiere un diagnstico diferencial con otras estructuras abdominales que se
pueden prolapsar a travs de heridas vaginales situadas ms cranealmente [33].
Las laceraciones y las contusiones que se pueden producir durante las fetotomas o la
extraccin de fetos enfisematosos, pueden motivar una severa celulitis perivaginal e
inflamacin plvica con abscedacin. Estas pueden ocasionar la muerte a vacas estresadas
e intoxicadas que no reciban un tratamiento adecuado [33].
Los desgarros del cuerpo del tero y del cuello pueden ser suturados con catgut a travs de
la vagina. Los de los cuernos, requieren prolapsar el tero o abordarlo mediante una
laparotoma en la regin inguinal o en la porcin caudal de la lnea media. El prolapso del
tero, durante las primeras horas del puerperio, se puede lograr con una inyeccin
endovenosa de 10 mL de epinefrina (1:1 000 USP), la que causa una relajacin completa
del miometrio.
La produccin del prolapso se puede facilitar, cogiendo un puado de carnculas situadas
lo ms prximas posible, al extremo del cuerno en que se desarroll la gestacin y
mediante una traccin suave, se sitan en el canal del parto. La presencia de las carnculas
en el canal, provoca contracciones peristlticas que producen el prolapso del cuerno. Este
procedimiento no debe ir precedido de anestesia epidural ni de aplicacin de oxitocina
[17].
Hemorragias
Siempre despus del parto hay alguna sangre fetal en la vulva, procedente de la ruptura del
cordn umbilical. Se debe hacer un examen manual para detectar hemorragias en sitios
ms internos [33]. Si sangra un vaso grande en la vagina se debe hacer una ligadura o
contener la hemorragia con una pinza hemosttica, la que debe quedar puesta por uno o dos
das.
Las heridas del cuello uterino originan hemorragias persistentes. Estas se producen cuando
se practica una extraccin forzada del feto, sin que la dilatacin cervical se haya
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
completado. El tratamiento se puede aplicar extrayendo el cuello con una pinza cervical,
despus de la aplicacin de anestesia epidural. Las hemorragias uterinas se pueden
disminuir con una aplicacin de 50 Ul de oxitocina, que puede contraer al tero y sus
vasos.
El color de las mucosas, el aumento de los movimientos respiratorios y del pulso, anuncian
el peligro de un choque hipovolmico. El valor del hematocrito y el estado de la paciente
indicarn la necesidad de una transfusin sangunea.
La ruptura de grandes vasos de los ligamentos anchos, ocasionan hemorragias fatales. La
ruptura de estos vasos por lo general se produce en casos de torsin uterina o de prolapso
uterino. Cuando se sospecha que existe esta hemorragia, la nica posibilidad para tratar de
salvar al animal es realizar una rpida laparatoma para ligar el vaso [17].
Otras medidas inmediatas con la madre
Si la vaca permanece echada ms de media hora despus del parto, se debe estimular para
que se pare, posicin que reduce la posibilidad del prolapso del tero. Adems esto asegura
que no haya fracturas, luxaciones o compresin nerviosa, que impidan al animal sostenerse
de pie, o "que no sufra una hipocalcemia.
Se deben ofrecer las condiciones necesarias al animal, para que no resbale cuando intente
pararse, y si es necesario ayudarlo sostenindolo por la cola.
La ubre se debe examinar para comprobar la existencia de calostro y detectar cuartos
ciegos o afectados de mastitis.
PUERPERIO
Por puerperio se entiende el perodo que transcurre desde el parto hasta que los rganos
genitales, y el estado general de la hembra vuelven al ordinario anteriora la gestacin.
Constituye un importante perodo en la vida reproductiva de las hembras, porque influye
enormemente sobre la fertilidad subsecuente de estas.
Dentro del perodo puerperal, tiene gran importancia el proceso de involucin uterina, el
cual garantiza la reduccin del rgano a su volumen inicial y, adems, propicia todos los
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
cambios necesarios para que el tero se prepare desde el punto de vista histolgico y
fisiolgico para recibir, anidar, nutrir y desarrollar el prximo embarazo. Este perodo ha
sido estudiado en la mayora de las especies domsticas, pero ha recibido una atencin
especial en la hembra bovina, por ser ella la de mayor inters econmico, por tal razn se
hace referencia al proceso de su involucin puerperal.
Las contracciones del tero mantienen a las 14 h despus del parto, la misma duracin y
frecuencia que en la expulsin de la placenta (8-10 contracciones cada 30 min).
Transcurridas las 24 o 48 h el ritmo regular desaparece y despus de las 96 h es posible
registrar algunas contracciones irregulares.
Las contracciones del miometrio disminuyen el volumen del tero longitudinal y
transversalmente, las fibras musculares del tero puerperal poseen la particularidad de que
despus de su contraccin la relajacin no es total, quedndose ms o menos retradas.
Solo as es posible el hecho que inmediatamente despus del parto el volumen del tero es
mucho menor, al que alcanz en el perodo de la gestacin.
Despus del parto el peso del tero es aproximadamente 10 Kg. y debe regresar una vez
transcurrida la involucin a su peso original (antes de la gestacin) de 1 Kg por lo que en
todo el proceso de involucin el tero debe reducir su peso en 9 Kg. Las observaciones
clnicas han demostrado que en la involucin normal los procesos durante los primeros
das posparto son muy rpidos.
Luego de los cuatro das el crvix puede estar parcial o totalmente cerrado y el peso del
tero puede ser de solo 3,5 Kg.; en igual perodo las grandes carnculas con un peso inicial
de 200 g al parto, se han aducido a 50 g. A los siete das posparto el tero con un peso de
ms o menos 2 kg, puede ser palpado con facilidad por va rectal y esto da la impresin de
una gestacin de 2,5-3 meses, y ya entre los 15 y 17 das se encuentra reducido casi al
tamao del tero vaco. Se debe aclarar que el tero siempre va a quedar algo mayor que
antes de la gestacin, es decir, la regresin total hasta el tero juvenil no es posible por los
cambios que se realizan durante el perodo evolutivo (hiperplasia del miometrio y del
tejido conjuntivo), de modo que la relacin entre el tero juvenil y el tero que ya tuvo una
gestacin es de 1:1,2 y esta relacin aumenta con el nmero de gestaciones.
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
Modificaciones muy importantes sufren tambin los vasos sanguneos, la arteria uterina
media reduce rpidamente su tamao y el frmito tpico de la preez disminuye durante las
primeras horas despus del parto, y desaparece completamente a los cuatro das, en el caso
de involucin uterina normal.
Los cambios histolgicos en la musculatura del tero tienen gran inters desde el punto de
vista del proceso de involucin uterina normal o fisiolgica, toda vez que inmediatamente
despus del parto aparece el glucgeno en la clula muscular del tero, sin embargo, a las
24 h se inicia la infiltracin grasa, la cual culmina a los ocho das despus del parto.
La infiltracin grasa se acompaa de una notable fragilidad del tero, la que se deber
tener presente, sobre todo, al examinar o tratar el tero puerperal. A partir de los ocho das
se realiza la reabsorcin de la grasa; las clulas disminuyen su tamao y despus de las 3 o
4 semanas se encuentran solamente restos de dicha infiltracin.
Uno de los momentos ms caractersticos del perodo puerperal es el flujo de los rganos
genitales con el que se eliminan del tero los productos nocivos de la involucin uterina;
esto ayuda en el proceso de la limpieza de la cavidad uterina. El flujo uterino (loquios)
durante el transcurso del puerperio vara en su composicin y caractersticas.
Inmediatamente despus del parto los loquios contienen parte de los lquidos fetales, restos
de la sangre placentaria y moco producido por el cuello uterino, ms tarde se enriquecen
con detritus de las carnculas (3-10 das posparto), clulas de descamacin endometrial,
glandulares, plasmticas, e histiocitos, linfocitos y escasos leucocitos.
Los loquios son mucosos y sanguinolentos en los primeros das despus del parto; luego de
los 3 o 4 das comienzan a observarse partculas del tejido caruncular degenerado, lo que
ocasiona la condensacin y el cambio de color de los loquios, que se tornan grisceos y
conservan el color rojo. Pasada una semana estos adquieren una consistencia ms pastosa
(densos) parecida al pus, lo que hace que aumente su densidad alrededor de los 12-14 das
posteriores al parto, con un color semejante al chocolate y es eliminado en muy poca
cantidad. Dentro de los 14-18 das del parto las secreciones se tornan transparentes, muy
parecidas al moco estral y desaparecen pocos das despus.
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
Una parte de los loquios se elimina al exterior, pero una gran cantidad se reabsorbe a travs
de la mucosa endometrial. As, el primer da siguiente al parto, el tero contiene de 1,4-1,6
L de lquido; posterior a los ocho das solamente 0,8-0,6 L y 14 das despus del parto, solo
queda un revestimiento en la superficie de la mucosa.
El perodo de la involucin uterina es muy variado entre las distintas especies, pero de
forma general se acepta que este proceso presenta una duracin ms prolongada en la vaca,
y que la rapidez en la regresin de los rganos genitales se encuentra altamente
influenciada por numerosos factores, tanto internos como externos, en especial la actividad
que presenten los ovarios; se ha comprobado que en hembras con actividad ovrica antes
de las tres semanas posparto, el proceso de involucin ha sido muy rpido (menos de 21
das).
En el caso de los pequeos rumiantes y la cerda, los loquios suelen fluir una semana o algo
ms. En la yegua el flujo- loquial es escaso y solo puede comprobarse durante algunos
das, se supone que en estas hembras la involucin uterina es muy rpida, pues en un
porcentaje elevado de hembras se presenta un celo de buena fertilidad a los 9-10 das de
haber parido. La perra elimina inmediatamente despus del parto -segn el nmero de
fetosun cogulo sanguneo mezclado con abundante moco y un pigmento verdoso
(pigmento placentario) procedente del hematoma marginal de la placenta. En las gatas este
pigmento es carmelita.
Reinicio de la actividad sexual posparto
En el transcurso de la gestacin, la sntesis de LH se halla inhibida por un feedback
negativo, producto de los altos niveles de progesterona; no obstante, su mecanismo de
liberacin se mantiene activo, por lo que sus depsitos en la hipfisis llegan a agotarse.
Despus de la expulsin del feto, comienza un proceso de involucin uterina que se
caracteriza por una elevada produccin de prostaglandina F2a, la que alcanza
concentraciones mximas del da 1 al 4 posteriores al parto, declinando gradualmente hasta
alcanzar niveles bsicos el da 15. Estos altos valores de prostaglandina F2a de los
primeros das del posparto deben ser los responsables de la rpida involucin del cuerpo
lteo de gestacin despus del parto.
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
La cada de los niveles de progesterona a que da lugar la involucin del cuerpo lteo de
gestacin, permite el restablecimiento de la sntesis de LH. Este se produce en dos fases, la
primera se desarrolla en las primeras semanas del puerperio y se caracteriza por pulsos de
GnRH de relativa poca frecuencia, que solo permiten pulsos de LH de pequea amplitud,
insuficientes para inducir la maduracin folicular. Esta fase se supone que es relativamente
independiente del estmulo del amamantamiento y de los factores estresantes del medio
[34].
La segunda fase comienza cuando los depsitos de LH de la hipfisis han retornado a la
normalidad y los pulsos de LH tienen suficiente amplitud para estimular el crecimiento
folicular. La frecuencia de descargas de GnRH aumentan, y con ello, la de los pulsos de
LH, que completan la maduracin folicular y producen la ovulacin. Esta segunda fase
parece estar fuertemente influida por el estmulo del amamantamiento y por los factores
estresantes del medio. El amamantamiento y los factores estresantes del medio, parece que
inducen la secrecin de pptidos opioides endgenos, los que inhiben las descargas de
GnRH del hipotlamo [34].
En la vaca productora de leche, que recibe una alimentacin adecuada y no amamanta a su
cra, la actividad ovrica comienza de 10 -14 das despus del parto y se produce una
ovulacin silente pasada una semana. Esta ovulacin da origen a un pequeo cuerpo lteo
de corta vida, que facilita el comienzo de ciclos normales alrededor de los das 25-30 del
posparto. No obstante, el primero de estos celos frecuentemente es dbil y se requiere el
auxilio de un toro para detectarlo.
Factores que influyen en el comienzo de la ciclicidad estral
El comienzo de la actividad cclica estral, est fuertemente influido por el nivel de
nutricin anterior y posterior al parto. Es bien conocido el efecto negativo que la prdida
de peso ejerce sobre la actividad ovrica. La vaca debe parir con una condicin corporal de
3,5, segn la escala establecida de 5 para que despus del parto quede en una de 3.
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
La lactacin es otro factor influyente, las vacas mastectomizadas entran rpidamente en
celo. Las que se ordean dos veces diarias, tienen intervalos ms cortos del parto al primer
celo que las que amamantan a un ternero, estas tienen un intervalo similar en duracin a las
que se ordean cuatro veces diarias. Las vacas que amamantan dos o ms terneros tienen
un intervalo de anestro mayor que las que amamantan uno solo.
El efecto de la raza es bien manifiesto en las vacas de las razas fos indicus, que
amamantan a su cra, las que tienen un anestro posparto mucho ms largo que el de las
vacas de las razas Bos taurus.
Las novillas que tienen su primer parto, invariablemente manifiestan un anestro posparto
mayor, que las vacas multparas.
INDUCCION DEL PARTO EN LA VACA
En 1969, Adams [35] desarroll un mtodo de induccin del parto en la vaca, utilizando
para ello 20 mg de dexametasona. De 22 hembras que trat, 19 parieron entre las 22 y
56 n de aplicado el tratamiento, con un. promedio de 45 h. De las 19 vacas que
parieron, 18 retuvieron la placenta, expulsndola solo una, cuya gestacin estaba a
trmino. Las tres que no respondieron al tratamiento tenan de 235-257 das de
gestacin.
Posteriormente, numerosos autores han reportado aumentos importantes de retenciones
placentarias cuando se induce el parto con glucocorticoides [36] [37] [38] [39] y [40].
Las placentas que son retenidas como consecuencia de los tratamientos de induccin del
parto, por lo general no pueden removerse sin causar considerables traumas [20], por lo
que est contraindicada su extraccin manual. Esta se ha comprobado que prolonga el
perodo parto primera ovulacin [41 ].
La aplicacin de potentes glucocorticoides puede alterar la frmula leucocitaria, con
aumento de neutrfilos y reduccin de los linfocitos circulantes, que persiste de 1-3 das
despus de una simple inyeccin. Adems, los animales tratados presentan un elevado
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
nivel de glucosa, como resultado de su actividad glucognica y una alteracin de su
respuesta inmunoigica [42].
La aplicacin de dexametasona de 8-14 das antes del parto esperado, produce una rpida y
significativa cada de la concentracin de progesterona en el plasma sanguneo, a niveles
similares al que se produce en el momento del parto [43].
En las vacas que se les induce el parto, el edema de la ubre es menor, por lo que se ha
recomendado su utilizacin, como tratamiento de las que presentan severo edema de la
ubre [44].
Durante la dcada del 70 se aplic la induccin del parto masivamente en Nueva Zelanda,
en dicha dcada se indujeron ms de un milln de partos prematuramente, con el propsito
de que los partos se produjeran al inicio de la primavera. Utilizaron para ello,
principalmente, corticoides insolubles de accin lenta, con el fin de disminuir el nmero de
retenciones placentarias. No obstante, las prdidas de terneros fueron muy cuantiosas, ya
que aproximadamente el 14 % nacieron muertos y la mortalidad neonatal fue de algo ms
del 17 % de los nacidos vivos [45].
Los glucocorticoides pueden ser administrados para inducir el parto en la vaca, la oveja y
la cabra, pero se ha visto que es menos efectivo en la yegua y la cerda [35].
Induccin del parto con prostaglandina
En 1974, Spears [46] indujeron el parto de 42 vacas con prostaglandina F2a, utilizando
para ello una dosis de 20 mg.
El parto puede ser iniciado por la administracin de prostaglandina en la vaca, la cerda, la
cabra y la yegua, as como en la mujer, no obstante, el mecanismo por el que induce el
parto tanto la prostaglandina F2a como sus anlogos vara: en la mujer es por la accin
directa sobre la actividad contrctil del miometrio y cambios cervicales, y en los citados
animales por induccin directa de lutelisis. Sin embargo, la prostaglandina F2a no induce
el parto en la oveja [3].
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
El primer reporte del uso de un anlogo de la prostaglandina F2a, fue en 1975, cuando
Bosh et al. [47] lograron inducir el parto con inyecciones repetidas de cloprostenol. Los
terneros procedentes de partos inducidos con prostaglandina tienen un vigor normal y la
induccin no causa unas muertas o muertes de recin nacidos. Estos al nacer muestran
menos peso [36] [48] y [49], pero crecen normalmente y ganan peso cuando maman al
mismo ritmo que los terneros controles.
El cloprostenol no produce un desarrollo muy rpido de la ubre, y la mayora de los
animales tratados con esta hormona, no muestran un desarrollo completo de esta hasta 4 o
5 das despus del parto, especialmente en aquellos casos que no presentan un ligero
desarrollo de la ubre el da del tratamiento [50].
A pesar de que el desarrollo de la ubre se produce entre el tratamiento de induccin y el
parto, la produccin de leche es menor a los seis das posparto, pero a los 20 das la
produccin de tratadas y no tratadas es similar (Tabla 5.2), al igual que la produccin total
[20] [37] [51] [52] y [53].
TABLA 5.2 PRODUCCIN DE LECHE ACUMULADA EN 35 VACAS DEL
PRIMER PARTO Y 35 CONTROLES (PROMEDIO EN kg) 130]
Das de lactancia
30
Partos inducidos
DS
CV
394,53
55,47
14,06
Partos no inducidos
X
DD
CV
391,24
71,27
17,94
60
792,53
102,25
12,90
783,41
125,21
15,98
90
1 179,72
137,73
11.67
1 153,48
155,95
13,52
La induccin del parto en las bfalas tampoco afecta la produccin de leche ni su
composicin [54].
El tratamiento con cloprostenol pudiera ser un mtodo efectivo para inducir el parto en los
casos de hidropesa de las membranas fetales [55] y es capaz de iniciar el proceso de
expulsin de un feto muerto o de uno momificado casos en que los glucocorticoides no han
mostrado efectividad [42].
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
Entre el 28 y el 100 % de las vacas y novillas inducidas con prostaglandina retienen la
placenta como mnimo durante 24 horas. El promedio de retencin es del 59 %. No se ha
encontrado diferencia en la retencin placentaria de vacas tratadas con prostaglandina F2a
o con Fenoprostalene combinado con dexametasona [56].
El grave problema de la retencin de la placenta restringi el uso de los corticosteroides y
de la prostaglandina en la induccin del parto en bovinos, solamente a los casos de
gestacin prolongada, patolgica y casos en que se sabe se pudiera producir una
desproporcin fetomaterna grande.
Es evidente que la retencin placentaria tena que ser resuelta, antes de que el
procedimiento de induccin del parto ganara universal aceptacin para obtener los partos
en das predeterminados, y para prevenir distocias en animales jvenes [57].
Induccin y sincronizacin de partos con lidocaina y prostaglandina
A partir del descubrimiento de Preval [58] de las propiedades de la Iidocaina, tanto en la
prevencin como en el tratamiento de la retencin placentaria en el bovino, se trabaj
intensamente en la bsqueda de un esquema de tratamiento que fuera capaz de inducir el
parto, que este se produjera en forma sincronizada, en horas del da, y que no tuviera las
dificultades con la retencin placentaria que impedan su generalizacin.
A fines de 1985 se logr un esquema de tratamiento con el producto anestsico local,
Iidocaina al 2,5 % con epinefrina (Labiofam) 40 mL y una pequea dosis de cloprostenol,
que aplicados por la va vaginal fueron capaces de sincronizar entre el 80 y 90 % de los
partos, y que estos se produjeran en horas del da. La aplicacin de 120 mL de la solucin
de Iidocaina al 2,5 % en el tero, en las dos primeras horas que siguen al parto permite
evitar los efectos indeseables que ocasionaba la retencin placentaria.
La aplicacin de la sincronizacin de partos permite obtener suero de recin nacidos libres
de inmunoglobulinas, con caractersticas similares al suero fetal, producto con un alto
precio en el mercado mundial. Este mtodo de obtencin tiene la ventaja de no requerir
sacrificio de animales, lo que disminuye sustancialmente su costo de produccin.
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
Otro gran beneficio que se obtiene con la sincronizacin de los partos es la de posibilitar su
atencin facultativa, con ello se reducen las cras muertas durante el parto (Tabla 5.3).
TABLA 5.3
CRAS
MUERTAS
EN
PARTOS
SINCRONIZADOS
Y NO
SINCRONIZADOS DE VACAS HOLSTEIN [59]
Partos
Cras muertas
Sincronizados
2 896
24
0,80
No sincronizados
8843
332
3,75
La sincronizacin de los partos permite, por primera vez en la historia de la enseanza de
la Obstetricia Veterinaria que los estudiantes puedan aprender esa materia atendiendo
dichos partos. Por ello, el Seminario sobre Reproduccin Animal, auspiciado por la
Fundacin Internacional Sueca, celebrado en Uruguay en abril de 1990, acord
recomendar la introduccin de la sincronizacin de partos en la enseanza de la obstetricia
en todas las escuelas de veterinaria.
Parto de la yegua
En la yegua, usualmente, 2 o 3 das antes del parto, se presenta salida de calostro con
aspecto parecido a cera.
El feto determina el da en que se produce el parto, pero la madre puede decidir a qu hora
se efecta este, ya que se ha observado en 501 partos de yeguas, que se produjo en el 86 %
de ellos, entre las 7:00 p.m. y las 7:00 a.m. con un pico de incidencia alrededor de la media
noche. Ellas tienen la habilidad de demorar el parto durante muchos das, cuando se
producen disturbios o si son constantemente observadas [14].
La yegua que va a parir, debe ser situada en un box limpio con una buena cama. Se le debe
lavar la vulva, el perin y la ubre y colocarle en la cola una venda limpia.
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
La fase de dilatacin en la yegua es de aproximadamente 1 h, aunque puede durar 2 o 3, y
se caracteriza por inquietud y dolor abdominal; el animal tiene tendencia a dar vueltas.
Mantiene la cola levantada y la mueve bruscamente, golpendose con ella el ano. Se mira
los costados e intenta cocearse el flanco.
La fase de expulsin de solo 10-15 min de duracin, comienza con contracciones
abdominales, que provocan la rotura del alantocorion, que se ve impedido de salir al
exterior detenido por sus inserciones placentarias. Su rotura se manifiesta por la salida a
borbotones de un chorro de lquido semejante a la orina, no muy abundante.
La continuacin de las contracciones se acompaa de la aparicin del amnios (de pared
ms gruesa), con relaciones menos estrictas, que le permiten sobresalir de la vulva, dejando
ver las extremidades del potro. El amnios al llegar al lmite de posibilidades de
exteriorizarse se rompe ante la presin de las extremidades fetales, lo que motiva la salida
de parte del lquido amnitico, que posee manifiestas propiedades lubricantes. La cabeza
fetal se introduce entre los labios vulvares y fuertes contracciones logran distender el anillo
vulvar y que el occipucio fetal salve ese obstculo [23]. Tras una breve pausa, con una
sucesin de esfuerzos expulsivos el trax pasa a travs de la vulva, seguido rpidamente
por las caderas.
La yegua, por lo general, pare echada y el potro al nacer permanece con las porciones
distales de sus extremidades posteriores en la vagina, conservando intacto el cordn
umbilical. Despus de 5-15 min, la yegua se mueve para pararse o el potro comienza a
moverse, las patas salen al exterior y el cordn se rompe a una distancia de 5-8 cm de la
pared abdominal. Durante este tiempo el potrico acumula sangre que estaba en la placenta
fetal, la que puede representar el 30 % de la sangre fetal [60]. Por tanto, no es aconsejable
en los partos asistidos, cortar el cordn umbilical inmediatamente de producirse el parto,
ya que esto puede representarle una prdida de ms de 1 L de sangre, lo que puede ser
crtico para la vida del recin nacido. La madre no debe ser molestada, ni debe romper el
cordn por lo menos hasta transcurridos 10 min despus del nacimiento.
Ocasionalmente la primera membrana en aparecer en la vulva es el alantocorion, rojo
oscuro. Esta membrana debe ser abierta para permitir al amnios, que contiene al feto, ser
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
expulsado primero [61]. Esto es necesario porque el alantocorion puede separarse por
completo de su insercin uterina y el potro asfixiarse.
Despus de la ruptura del cordn umbilical se le debe hacer una aplicacin de yodo al
ombligo y se le debe extraer de 300-500 mL de calostro a la madre y suministrrsela al
potro. Si a la yegua no se le haba aplicado toxoide tetnico antes del parto, se le debe
administrar antitoxina tetnica al potro.
Parto de la oveja
El parto de la oveja, generalmente, est precedido de un descenso de la temperatura rectal
de 0,5 C, que la hace caer por debajo de 39,2 C, lo que ha sido propuesto como mtodo
para seleccionar las ovejas que van a parir en un trmino de 48 h, con una efectividad del
80 % [62].
El parto en esta especie es bastante parecido al de la bovina; las contracciones uterinas
comienzan alrededor de 12 h antes del parto y causan la dilatacin del cuello uterino. En
ocasiones 2-8 horas antes de la expulsin del primer feto las contracciones y las
relajaciones del tero se suceden sin pausa y su amplitud se incrementa [11]. Luego de
producirse la dilatacin cervical y concluir con esta la fase de dilatacin, los receptores del
cerviz y la vagina son activados y al penentrar el feto en esta ltima se originan descargas
de oxitocina que aumentan la contractilidad uterina [63]. En ese momento empiezan
contracciones abdominales sincronizadas con las ondas de contracciones uterinas que
provocan la expulsin del feto en unos 15min [1].
La pausa entre la expulsin de un feto y otros posibles fetos, oscila entre unos minutos y
dos horas [24]. El nacimiento espontneo del corderito a diferencia del de los temeros se
puede producir en actitud de flexin de encuentro bilateral. Posterior a la expulsin del
feto, las contracciones uterinas continan con igual intensidad, para expulsar la placenta
que en la mayora de los casos tiene lugar dentro de las 2 h siguientes.
Parto de la cerda
La gestacin de la cerda dura alrededor de 114-116 das, con un rango de 110-120. Al
parecer se requiere la cada de los niveles de progesterona circulantes para que se inicie el
parto.
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
Durante los ltimos das de la gestacin usualmente se aprecia una pronunciada
inflamacin de la vulva, con una coloracin que se hace ms roja cuando el parto se
aproxima. Cuando fluye la leche por las mamas, el parto se produce en un trmino de 6-12
h, aunque, en ocasiones, este puede demorar hasta 24 h. El da que precede al parto la
cerda se muestra nerviosa, intranquila, se echa y se levanta repetidamente y comienza a
preparar la cama. Cuando se va aproximando este, la respiracin se acelera, y defeca y
orina con frecuencia, se va tranquilizando y ms calmada se echa y generalmente pare
echada sobre un costado, con solo ocasionales cambios a una posicin ventral, o a pararse.
El incremento de las contracciones uterinas se manifiesta por movimientos abdominales y
de las patas y por el derrame de pequeas cantidades de fluido a travs de la vulva.
Antes del primer nacimiento pueden romperse varios sacos corinicos prximos al cuello,
los que proveen un tnel resbaladizo por el que se deslizan los lechones [60]. Los cordones
umbilicales de las primeras cras permanecen intactos y son lo suficientemente largos para
permitir que estas sean paridas antes de romperse, pero cuando el parto progresa muchos se
rompen, igualndose hacia los finales del parto, el nmero de cordones sanos y rotos.
El paso de cada lechn por la vagina con frecuencia es sealado por un movimiento de la
cola [64], su expulsin solo requiere una pequea contraccin de la madre, tanto la de los
que se encuentran en presentacin anterior como posterior. Algo ms de la mitad de los
cerditos nacen en presentacin ceflica.
El perodo de expulsin en la cerda dura de 1-4 h, pero puede llegar a 8 h. El intervalo
entre el nacimiento de una cra y otra es alrededor de 15 min, aunque pueden ser paridas
rpidamente o presentar retrasos entre unas y otras. Estas pausas entre un nacimiento y
otro, casi siempre son menores entre los primeros nacimientos y mayores entre los ltimos.
En cuanto al orden de los nacimientos, el primero en nacer es el que est ms prximo al
cuello uterino, pero despus los nacimientos se producen sin guardar ningn orden, se
puede alternar, la salida de los que ocupan los cuernos derecho e izquierdo o pueden salir
primero todos o casi todos los de un cuerno.
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
Al nacer el recin nacido permanece inmvil por unos segundos. Cuando comienza la
actividad respiratoria trata inmediatamente de liberarse de la atadura que representa el
cordn umbilical, si este no se haba roto, y una vez libre avanza hacia adelante en
direccin a la ubre.
En ocasiones un parto se considera terminado y un da e incluso dos das ms tarde se
produce el nacimiento de otro cerdito vivo.
Expulsin de las membranas fetales
Por lo general las placentas se expulsan al final del parto, despus de las cras, aunque la
ltima de estas puede ser expulsada durante o entre la expulsin de las placentas. En
ocasiones varios (echones pueden ser seguidos de varias placentas.
Cras muertas
La mayora de las cras muertas (alrededor del 80 %) se producen en el ltimo tercio de la
gestacin. La causa de estas muertes por anoxia que sufren esos cerditos, es la posicin
ms distante del crvix que ellas ocupan en el tero. Como consecuencia de la mayor
distancia a recorrer hay mayor cantidad de casos de ruptura del cordn umbilical. Adems,
estas cras son las ltimas en nacer, encontrando un tero fatigado que hace ms lento el
proceso de expulsin. Se ha observado que las pausas entre el nacimiento de una y otra cra
son mayores cuando preceden a una cra muerta, no estando completamente claro si la
pausa es motivo o consecuencia de la muerte fetal [64].
Asistencia al parto
Generalmente los partos de la cerda solo requieren la vigilancia del proceso de expulsin,
para liberar a las cras de envolturas fetales que puedan impedir su respiracin, cuidar que
la madre no aplaste a las cras o se las coma y separar los cerditos muertos y las placentas
expulsadas.
Parto de la perra
El parto de la perra se produce ms o menos cerca de los 63 das despus de la ovulacin o
aproximadamente 57 das despus del cambio de la citologa del frotis vaginal, de un
predominio de clulas cornificadas a predominio de clulas no cornificadas, cambio que
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
marca el primer da del diestro [65]. Debido al largo perodo del estro, la gran variacin
entre el comienzo de este y la ovulacin y la larga supervivencia del esperma de perro en el
tero de la perra, cuando estas hembras conciben despus de una simple monta, pueden
parir en un rango que va de los 58 a los 71 das posteriores a dicha monta.
Signos de la proximidad del parto
Al aproximarse el parto hay un rpido incremento de peso con marcado aumento de
volumen del abdomen y de las mamas, as como edematizacin de la vulva. Cuando el
parto es inminente, hay relajacin de los ligamentos plvicos, cambios en el
comportamiento, e intentos de preparar su cama. En las primigrvidas la lactacin coincide
con el parto, pero en las multigrvidas se puede extraer leche de las mamas desde varios
das antes de que este se produzca.
Una sbita y transitoria cada de la temperatura de alrededor de un grado centgrado es
usual que se produzca dentro de las 24 h anteriores al parto, la que se ha relacionado con la
cada del nivel de progesterona por debajo de 2 ng/mL.
Primera fase del parto
La primera fase del parto, fase de dilatacin, puede durar de 6-8 h y llegar a 12 h en las
primigrvidas. En esta fase la perra presenta anorexia, se muestra inquieta, tiembla y, en
ocasiones, puede vomitar, trata de esconderse y acondiciona su cama, aunque algunas
prefieren la compaa de su amo. Como algunos de estos signos se pueden presentar
intermitentemente en la ltima semana de la gestacin, puede ser difcil para el dueo
saber cundo comienza esta primera fase del parto y como consecuencia su duracin [65].
Segunda fase del parto
El comienzo de la segunda fase, fase de expulsin, est marcado por el inicio de los
esfuerzos expulsivos. El animal lame a menudo su vulva, lo que anuncia la prxima salida
del feto. La rotura de la primera bolsa de las aguas, alantoides, se puede producir dentro de
la vagina, o aparecer en la vulva como una pelota del golf [23], la que puede ser rota por la
madre con los dientes. La salida de la cabeza del feto requiere como en otras especies los
mayores esfuerzos, pero el resto del cuerpo casi siempre sale sin dificultad.
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
Cada cachorro nace cubierto por su amnios, pero la madre lo lame hasta liberarlo, limpiarlo
y estimular sus funciones respiratoria y cardiovascular [9]. El cordn umbilical es cortado
por su madre con sus dientes, tanto la seccin por mordedura, como el desgarre natural del
cordn, cohben con eficacia la hemorragia. Aproximadamente el 60 % de los cachorros
nacen en presentacin anterior o ceflica y el 40 % restante lo hace en presentacin
posterior o caudal.
La expulsin del primer teto se puede producir en solo 15 min, pero puede requerir hasta
una hora. Despus de su expulsin la madre tiene un descanso en el que permanece echada
y lame las secreciones de su vulva. De 1/2 a 2 h ms tarde, comienzan de nuevo los
esfuerzos expulsivos. La expulsin de los cachorros es muy irregular, aunque algunas
perras van pariendo a intervalos regulares. La duracin total del parto en las grandes
carnadas generalmente no rebasa 8 o 12 h, en algunos casos solo 4 h o aun menos. El parto
de la perra se caracteriza por secreciones de color verde oscuro, debidas a que contienen un
pigmento que procede del hematoma marginal de sus placentas y es liberado al
desprenderse estas.
Tercera fase del parto
La tercera fase o alumbramiento se produce tambin de forma irregular, en algunos casos
las membranas fetales son expulsadas en 10-15 min despus de cada feto y tambin puede
nacer un feto con las membranas de su predecesor alrededor del cuello, en tanto las propias
vienen detrs arrastradas por el cordn umbilical [23]. Normalmente la madre se come las
membranas fetales despus de su expulsin y en ocasiones pueden producirle vmitos [9].
Asistencia al parto
La atencin facultativa del parto de la perra, solo se requiere cuando el paso del primer feto
a travs de la vulva es dificultoso; se recomienda hacer traccin cuidadosa de dos pliegues
cutneos laterales del cuello o en la regin lateral del abdomen [24]. Para la asistencia del
parto de un feto en presentacin posterior, se recomienda como en todas las especies,
acelerar el parto despus de la salida del tren posterior, mediante una traccin rpida, para
evitar que el feto aspire lquido amnitico si Su cordn umbilical es comprimido.
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
Parto de la gata
Dos das antes del parto la gata se muestra muy nerviosa, algunas veces anorxica y
comienza a preparar su nido. Las mamas se hallan inflamadas y los pezones turgentes.
La hembra comienza el parto echada lateralmente hasta que las contracciones se hacen
intensas. Cuando el feto es empujado al canal pelviano cambia su posicin y se acuclilla.
Ella puede maullar cuando puja para expulsar su primer gatito, 5-15 min despus tras
rpidas contracciones expulsa la placenta que la madre come. Es frecuente que los dos
primeros gatitos sean expulsados, uno tras otro. Una espera de 10-90 min, puede anteceder
a la expulsin del tercer gatito. El parto puede durar de 2-6 h.
Algunas gatas tras la entrega de las primeras cras puede tener un descanso de 12-24 h.
Durante este tiempo ella amamanta a estas y despus contina el parto [66].
Las carnadas, por lo general, son de 4-6 gatitos, pero en el primer parto pueden tener
carnadas ms pequeas. Estos abren sus ojos en el trancurso de la segunda semana de
nacidos.
c.- PATOLOGIAS DEL PARTO Y PUERPERIO
OBJETIVO
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
El estudiante con criterio cientfico-tcnico podr diagnosticar, controlar, tratar
eficientemente las diferentes patologas del parto y puerperio que se presente en la
vida cotidiana.
c.- Exploracin tocologica y preparativos para la asistencia al parto patolgico
1.- Intervencin al parto patolgico
2.- Anomalas de los dolores del parto
- Inercia Uterina primaria y secundaria
3.- Insuficiencias en la dilatacin del cuello uterino
4.- Cistocele vaginal
5.- Distocias fetales
- Distocias dependientes del tamao fetal
- Distocias debidas a actitudes defectuosas
- Distocias dependientes de la posicin
- Distocias por desproporciones feto- plvicas
- Distocias debidas a monstruosidades fetales
6.- Tokurgia. Fetotomia
7.- Operacin Cesaria
8.- Prolapso Uterino
9.- Retencin Placentaria
10.-Sndrome de la vaca caida
11.- Metritis Puerperal Septica
Con este acpite los estudiantes realizarn investigacin extracurricular mediante revisin
de literatura actualizada y tendrn que exponer la temtica designada.
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
REFERENCIAS BIBLIOGRFICAS
1). Me DNALO, L.E.: Veterinary endocrnology and reproduction. 3th ed., Lea and
Febiger, Philadelphia, 1980.
2). LANMAN, J.T.: Parturition in nonhuman primates. Biology of reproduction, 16:2838,1977.
3). MALTIER, J.P.; CH. LEGRAND and M. BREVILLER: Parturition. En:
Reproduction in mammals and man. (Eds.) C. Thibault; M.C. Levasseur and R.H.F.
Hunter, Ellipses, Pars, 1993.
4). UMO, I.; R.J. FITZPATRICK and W.R. WARD: Parturition in the goat: plasma
concentrations of prostaglandin F and steroid hormones and uterine activity during
late pregnancy and parturition. J. Endocrinol., 68: 383, 1976.
5). LIGGIN, G.C.; S.A. GRIEVES; S.F. KENDALL and B.S. KNOX: The physiological
role of progesterone, oestradiol-17 and prostaglandin 2 in the control of ovine
parturition. J. Reprod. Frtil., 16: 85,1972.
6). SPEARS, L.L.; B. BERCOVITZ; W.L. REYNOLDS; J.L. KREIDER and R.A.
GODKE: Induction of parturition in beef cattle with estradiol and PGF2a. J. Anim.
Sci., 39(1): 227,1974.
7). PETERSON, A.J.; J.T. HUNTER; R.A.S. WELCH and R.J. FAIRCLOUGH:
Oestrogens in bovine fetal and maternal plasma near term. J. Reprod. FertH.,
43:179,1975.
8). KNICKERBOCKER, U.; M.C. WILTBANK and G.D. NISWENDER: Mechanism
ot luteolysis in domestic livestock. Domest. Anm. Endocrinol., 5: 91, 1988.
9). BENNET, D.: Normal and abnormal parturition. En: Current therapy in
theriogenology. (Ed.) D.A. Morrow, W.B. Saunders Company, Philadelphia, London,
Toronto, 1980.
10). MARTN MARTN, E.: Fisiopatologa de la reproduccin con sus bases sinpticas.
Instituto Experimental de Ciruga y Reproduccin de la Universidad de Zaragoza,
Espaa, 1985.
11). FITZPATRICK, R.J.; Pregnancy and parturition. En: Current therapy in
theriogenology 2. (Ed.) D.A. Morrow, W. B. Saunders Company, Philadelphia,
London, Toronto, 1986.
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
12). BELT, W.D.; L.L. ANDERSON; L.F. GABAZOS and R.M. MELAMPY:
Cytoplasmic granules and relaxin levis in porcine corpora ltea. Endocrinology,
89:1,1971.
13). SHERWOOD. O.D.: Relaxin at parturition in the pig. En: Control of pig
reproducton. (Eds.) D.J.A yG.R. Foxcroft, Butterworths, London, 1982.
14). BAZER, F.W. and N.L. FIRST: Pregnancy and parturition. J. Anim. Sci., 57 sup. 2:
425-460,1983.
15). SHMIDT y ELLENDORFF: Endocrinologa y fisiologa de la reproduccin de los
animales zootcnicos. Ed. Acribia, Zaragoza.1972.
16). SLOSS, V. and J.H. DUFTV: Manual de obstetricia bovina. Compaa Editorial
Continental, S.A. de S.V. Mxico, 1987.
17). MARTEN, D.: Periparturient care of the dam. En: Current teherapy in
theriogenology. (Ed.) D.A. Morrow, W. B. Saunders Company, Philadelphia, London,
Toronto, 1980
18). ROBERTS, S.J.: Veterinary obstetrics and genital diseases. 2nd. ed., Edwards Brothers
Inc. Ann. Arbor, Michigan, 1971.
19). WAGNER, W.C.: Parturition induction in cattle. En: Current therapy in
theriogenology. (Ed.) D.A. Morrow, W.B. Saunders Company, Philadelphia, London,
Toronto, 1980.
20). WILLIAMS, W.F.; M.J. MARGOLIS; J. MANSPEAKER; L.W. DOUGLAS and J.R
DAVIDSON: Peripartum changes in the bovine placenta related to fetal membrane
retention. Theriogenology, 28(2): 213-223,1987.
21). WINFIELD, C.G. and A.W. MAKIN: Prediction of the onset of parturition in sheep
from observations of rectal temperature changes. LivestockProd. Sci., 2:393,1975.
22). GRUNERT, E.: Etiology of retained bovine placenta. En: Current therapy in
theriogenology. (Ed.) D.A. Morrow, W. B. Saunders Company, Phila-delphia, London,
Toronto, 1980.
23). ARTHUR, G.H.; D.E. NOAKES and H. PEARSON: Veternary reproduction and
obstetrics, 6th. ed., Bailliere Tindall, London, 1989.
24). BENESCH, R: Obstetricia y ginecologa veterinarias. Editorial Labor, S.A., Madrid,
1963.
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
25). SHUIJT, G. and L. BALL: Delivery by torced extraction and other aspects of bovine
obstetrics. En: Current therapy in theriogenology. (Ed.) D.A. Morrow, W.B. Saunders
Company, Philadelphia, London, Toronto, 1980.
26). HINDSON, J.C.: Quantification of obstetric traction. The Veternary Record,
102:327-332,1976.
27). MAARTEN, D.: Perinatal care of the call. En: Current therapy in theriogenology.
(Ed.) D.A. Morrow, W. B. Saunders Company, Philadelphia, London, Toronto, 1980.
28). TORRES, OLGA; MARISELY GONZLEZ y J.M. CAPOTE: Efecto del parto
dificultoso sobre el estado cido-bsico efel ternero neonato. Rev. Salud Anim.,
7:255-260,1985.
29). TORRES, OLGA: Sistema de atencin al parto y al ternero recin nacido, Folleto del
CENSA, La Habana, 1988.
30). MAARTEN, D.: Principies of calostrum feeding. En: Current therapy in
theriogenology. (Ed.) D.A. Morrow/W. B. Saunders Company, Philadelphia, London,
Toronto, 1980.
31). BRUSON, D.B. and J. W. LUDDERS: Resucitation and intesive care of the newborn
calf. En: Current therapy in theriogenology 2. (Ed.) D.A. Morrow, W.B. Saunders
Company, Phlladelphia, London, Toronto, -1986.
32). LE BLANE MICHELLE, M..: Passive transfer of inmunity in calves. En: Cyrrent
therapy in
theriogenology 2. (Ed,) D.A. Morrow, W. B Saunders. Company,
Philadelphia, London, Toronto, 1.986.
33). HAIBEL, G.K.: Inmediato postpartum care of, tne dam>>. Efr,Cunenttherpy in
theriogenology 2. (Ed.) D.A. Morrow, Wt, B. Saunders; Gomparjy, Philadelphia,
London, Toronto, 1986.
34). NETT, TX; Function of the hipotalamic-hipofisial axis during the postpartum period
in ewes and cows. J. Reprod. Frtil., suppl. 34, 200-203,1987.
35). ADAMS, W. M.: The elective induction of labor and parturition in cattle, J. Amer.
Vet. Med. Ass-, 154: 261-265, 1969.
36). BEARDSLEY, G.L; L.D. MULLER; M.J. OWENS; FL.C. LUDENS and W.L
TUCKER: Initiation of parturition in dairy coros with dexamethasone I cow response
and performance. J. DairySci-, 57(9): 1061-1066,1974.
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
37). BOLTE, K.A.; H.A. GARVERICK; D.J. KESLER; B.M. DAY and E.C. MARTHER:
Dexamethasone and estradiol benzoate induced parturition in dairy cattle.
Theriogenology, 8(1): 45-48, 1977.
38). CHEW, B.R; R.E. ERB; R.D. RANDEL and F.M. ROQUETTE: Efecct of corticoid
induced parturition on lactation and on prepartum pretiles of serum progesterone and
the estrogens among cows retaining and not retaining fetal membranes.
Theriogenology, 10(1): 13-25,1978.
39). GROSS, T.S.; W.F. WILLIAMS and T.W. MORELAND: Prevention of the retained
fetal membrane syndrome (retained placenta) during induced calving in dairy cattle.
Theriogenology, 27(3): 365-369,1985.
40). MORRISON, D.G.; PE. HUMES and R.A. GUDKES: The use of dimenhidrinate in
conjunction with Dexamethasone for induction of parturition in beef cattle.
Theriogenolgy, 19(2): 221-223, 1983.
41). BOLINDER, A.; B. SEGUIN; H. KINDAHL; D. BOULEY and D. QTTERBY:
Retained fetal membranes in cows. Manual removal versus non removal its effects
on reproductive performance. Theriogenology, 30(1): 45-46,1988.
42). WAGNER, W.C.; R,L. WILLHAM and L.E. EVANS: Controlled parturition in
cattle. J. Anim. Sci., 38(3): 485-489,1974.
43). GARVERICK, M.A.; B.N. DAY; E.C. MATHER; L. GMEZ and Y. THOMPSON:
Use of estrogen with dexamethasone for inducing parturition in beef cattle. J. Anim.
Sci., 38(3): 584-590,1974.
44). SCHMITT, D.; H.A. GARVERICK; E.C. MATHER; J.D. SIKES; B.N. DAY and R.E.
ERB: Induction of parturition in dairy cattle with dexamelhasone and estradiol
benzoato. J. Anim. Se.., 40(2): 261-268, 1975.
45). MOLLER, K. and S.C. MAC DARMIO: The economy of induced early parturition
in New Zeland dairy herds. A/. Z. Vet. J., 29(9): 165-167, 1981.
46). THORBUN, G.D. J.R. CHALLIS and W. BRUCE CURRIE: Control of parturition
in domestic animals, Biology of Reproduction, 16(1): 18-27, 1977.
47). Bosc, M.J.; J. FEURE and VASLET DE FONTAUBERT: A comparison of
indruction of parturition with dexamethasone or with an analog of prostaglandin F in
cattle. Theriogenology, 3(5): 187-191,1975.
48). BEARDSLEY, G.L.; L.D. MULLER; H.A. GARVERICK; F.C. LuDENSvand W.L.
STUCKER: Initiation of parturition in dairy cows with dexamethasone. II Response
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
to dexamethasone in combination with estradiol benzoate. J. Dairy Sci, 59(2): 241
-247,1976.
49). BERMOND, D.: Induccin del parto en la hembra bovina. Trabajo de Diploma,
Facultad de Medicina Veterinaria, ISCAH, La Habana, 1987.
50). DAY, A.M.: Cloprostenol for termination of pregnancy in cattle. a) The induction of
parturition. N.Z. Vet. J., 25(6): 136-139,1977.
51). JOHNSON, C.T.: Induction of parturition in cattle. Acta Vet. Scand., suppl. 77,311319.1981.
52). MNDEZ, MARGARITA: Estudio comparativo del comportamiento productivo y
reproductivo de vacas Holstein con partos naturales e inducidos. Trabajo de Diploma,
Facultad de Ingeniera Pecuaria, ISCAH, 1987.
53). O'FARREL, K.J.: Betamethasone induced calving: A comparison betwen induced
and not induced dairy cows. Calving problems and early viability of the call. Current
Topics in Vet. Med. and Anim. Sci., 4:341-351, 1979.
54). PRAKASH, B.S. and M.L. MADAN: Induction of parturition in water buffaloes
(Bubalusbubalus). Theriogeology, 23 (2): 325-331,1985.
55). SHARP, A.J.; C.J. BIERSCHWALL; R.G LMORE; R.S. YOUNQUIST; Al.
JENKINS; D.J. KESLER and H.A. GARVERICK: Response of two cows with
Cloprostenol (ICI80 996). Theriogenology, 10(1): 27-33,1978.
56). ECHTERNKAMP, S.E.; W.G. HAYS and W.G. KVASNICKA: Synchronization of
parturition in beef cattle with prostaglandin and dexamethasone. Theriogenology,
28(3): 227-247,1987.
57). KORDST, E. and W. JOCHLE: Induced parturition in dairy cattle. A comparison of a
corticoid (flumethazone) and e prostaglandin (PGF2a) in diferent age groups.
Theriogenology, 3(5): 171-178,1975.
58). PREVAL, B. y R. BRITO: Empleo del anestsico local (Lidocana al 2.5 % con
epinefrina) en el tratamiento de la retencin placentaria en la vaca. Informe Tcnico.
Dpto. Reproduccin, Centro Nacional de Salud Animal ISCAH, 1983.
59). BRITO, R. y B. PREVAL: Induccin del parto en la vaca, fes. Cub. Cien. Vet.,
20(4): 219-222,1989.
60). MC FDONALD, LD Endocrinologa veterinaria y reproduccin. 4ta. Ed., Nueva
Editorial Interamericana Me. Graw- Hill, Mxico, 1991.
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
61). HILLMAN, R.B.: Induction of parturition in the mare. En: Current Therapy in
theriogenology, (Ed.) D.A. Morrow, W. B. Saunders Company, Philadelphia, London,
Toronto, 1980.
62). WOOOING, F.B.R: Frecuency and localization of binucleated cells in the
placentomes of ruminants. Placenta, 4:527-540, 1983.
63). HINDSON, J.C.; B.M. SCHUFIELD; C.B. TURNER and H.S. WOLFF: Parturition in
sheep. J. Phisiol., 181:560,1965.
64). DAY, B.N.: Parturition. En: Current therapy in theriogenology. (Ed.) D.A. Morrow,
W. B. Saunders Company, Philadelphia, London, Toronto, 1980.
65). SHIRLEY D., JOHNSTON: Parturition and dystocia in the bitch. En; Current
therapy in theriogenology 2. (Ed.) D,A. Morrow, W.B. Saunders Company,
Philadelphia, London, Toronto, 1986.
66). CHAFFAUC, S. Reproduction of the eat and the dog. En Reproduction in mammals
and man. (eds). C. Thibault; MC. Levasseur and R.H.F. Hunter, Ellipses, Paris, 1993.
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
6. BIOTECNOLOGA DE LA REPRODUCCIN
El impetuoso avance de la biotecnologa obliga a ampliar cada da ms el contenido
cientfico de la reproduccin animal.
Irrumpi la transferencia de embriones en la mejora gentica, ampliando la limitada
participacin que la hembra tena en ella. Mediante blastmeros totipotentes abri el
camino a increbles resultados con el empleo de clulas diferenciadas (oveja Dolly).
El dominio de la manipulacin del genoma, capaz de modificar por primera vez, la
codificacin gentica, preocupa hoy a la humanidad ante la posibilidad de poner en manos
del hombre el porvenir de las especies y el del propio hombre.
Transferencia de embriones
La transferencia de embriones constituye un mtodo de obtencin de vulos fertilizados de
una hembra (donante) y su transferencia al aparato reproductivo de otra (receptora) de la
misma especie, donde se desarrolla la gestacin y se produce el parto.
Por su complejidad y limitaciones tcnicas y fisiolgicas la transferencia de embriones no
puede aspirar a reemplazar a la inseminacin artificial. No obstante, sus propiedades
utilizadas con animales seleccionados puede ofrecer un medio de progreso a travs de la
mayor utilizacin del mejor material gentico [1].
Esta tcnica facilita:
1.
Aprovechar al mximo el genofondo y el potencial reproductivo de hembras muy
valiosas.
2.
Utilizar los vientres de animales sanos y fuertes de escaso valor gentico.
3.
Intercambiar material gentico internacionalmente.
4.
Introducir rpidamente una raza no existente en un pas.
5.
Transportar hatos enteros en forma de embriones congelados, con un costo inferior al
costo del transporte de un solo animal adulto.
6.
Obtener ms de una descendencia de cada embrin, con lo que se logran animales
genticamente idnticos (similares a los gemelos unvitelinos) que posibilitan realizar
importantes trabajos de investigacin cientfica.
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
7.
Aprovechar como donantes vacas valiosas que sufren leucosis u otras enfermedades
infecciosas, que mediante receptoras sanas permiten obtener cras sanas de alta
calidad, dado que es muy improbable que el embrin transmita enfermedades
infecciosas.
8.
Formar hatos de vacas lecheras libres de leucosis bovina.
9.
Obtener vulos de terneras hijas de padres de alto valor gentico, para lograr
descendientes de estas antes de que realicen su primer parto, abreviando de esta forma
su intervalo de generacin.
La transferencia de embriones, en la actualidad es aplicada a casi todas las especies de
animales domsticos y a muchas especies de animales salvajes y exticos e inclusive a la
especie humana.
La primera transferecnia de embriones la realiz Heape en conejas en 1890 y el primer
embrin de bovinos se obtuvo en 1930. No obstante, no fue hasta principios de la dcada
del 70 que se despert el inters comercial en la transferencia. En Cuba, en 1976, el Dr.
Carlos Iglesias Oliva obtuvo los dos primeros nacimientos de temeros, llevando a cabo esta
tcnica por la va quirrgica (decimoquinto pas en el mundo y primero en Amrica Latina)
y en 1984 se lograron ocho cras de la recordista mundial Ubre Blanca. En el mismo ao se
obtienen las primeras gestaciones con embriones congelados y en 1985 una gestacin con
un hemiembrin. Hoy da se cuenta con una red de 12 centros, distribuidos en todas las
provincias del pas.
En agosto de 1992 naci el primer ternero por fecundacin in vitro, siendo Cuba el primer
pas en Amrica Latina y el duodcimo en el mundo en lograrlo.
Seleccin de las donantes
La aptitud de un animal como donante para la transferencia de embriones se determina por
su valor gentico y su capacidad para lograr un alto nivel de resultados. Estos pueden ser
ptimos si se puede seleccionar un animal que ya ha ofrecido buenos resultados en la
transferencia. Se debe tener en cuenta que un tercio de las vacas donantes no producen
embriones transferibles.
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
Se ha sugerido que en la seleccin de la donante se utilicen los criterios siguientes [2]:
1.
Haber presentado ciclos regulares desde temprana edad.
2.
No requerir ms de dos servicios por concepcin.
3.
Haber tenido tres cras dentro de dos aos calendarios.
4.
No haber tenido partos distcicos ni irregularidades reproductivas.
5.
No presentar defectos de conformacin o genticos detectables.
6.
Tener de 3-10 aos de edad.
Seleccin de las receptoras
Como receptora de un embrin de alta calidad gentico-productiva, se debe seleccionar un
animal sano con las mayores posibilidades de desarrollar su gestacin, por lo que es
conveniente destinar a este fin novillas con buen desarrollo corporal (340-350 kg) con
ciclos estrales regulares sometidas a un plano incrementado de nutricin.
Las novillas que repiten el celo despus de la transferencia, se le repetir el proceso una
vez ms, pero si vuelven a repetir sern inseminadas y se prescindir de ellas.
En un hato con buen manejo las vacas pueden ser tan buenas como las novillas, o mejores,
por tener menor mortalidad perinatal [3]. En nuestras condiciones de clima clido y
hmedo hay que tener presente que las vacas Holstein sufren mucha mortalidad de
embriones, especialmente en los meses clidos.
Si se utilizan vacas, estas deben ser jvenes y esperar que transcurra su segundo celo
pospartal, las que repitan el celo despus de embrionizadas sern inseminadas en ese
mismo celo [4].
Tcnica de transferencia de embriones
La aplicacin de la tcnica de transferencia de embriones requiere las operaciones
siguientes:
1. Superovulacin de la donante.
2. Sincronizacin de los celos de la donante y la receptora.
3. Inseminacin de la donante.
4. Recoleccin de embriones.
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
5. Seleccin de embriones.
6. implantacin de embriones.
Superovulacin de la donante
La aplicacin de un tratamiento superovulatorio a la donante, tiene como objetivo
aumentar el nmero de embriones frtiles normales, que permita una explotacin adecuada
de la transferencia de embriones [5].
El tratamiento superovulatorio es un rea que requiere estricta atencin, porque de esto
dependen los resultados que se obtengan con la transferencia de embriones. A pesar de ello,
las investigaciones realizadas en la dcada pasada no han provisto la informacin requerida
para reducir la variabilidad en la respuesta a los tratamientos superovulatorios, incrementar
el porcentaje de fertilizacin o aumentar la calidad de los embriones [6].
La variabilidad de la respuesta ovrica es muy grande; el 30% de las donantes no producen
embriones transferibles y solo el 45% produce cuatro o ms embriones con un rango de 047 [1]. La causa de esta variabilidad depende, en cierto modo, de la existencia de diferentes
nmeros de folculos y de su Sensibilidad, pero tambin de la efectividad de las
preparaciones hormonales que se utilicen.
Con la tecnologa existente actualmente, el promedio para cada vaca donante superovulada
puede ser de 8-10 huevos colectados, de 6-7 embriones transferidos y 3-4 gestaciones [2],
pero en nuestras condiciones se obtienen resultados ms pobres, probablemente debido a
las condiciones climticas.
FACTORES QUE INFLUYEN EN LA SUPEROVULACIN
Los resultados de los tratamientos superovulatorios son muy variables, ya que se ven
influidos por muchos factores que pueden ser, la raza, la edad, la categora, la poca, el
nivel de nutricin, inmunolgicos e inclusive individuales.
Raza. Existe una notable diferencia a la respuesta hormonal entre razas [7] probablemente
debido a diferencias en el nmero de folculos. La raza Charolaise da una respuesta mayor
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
a los tratamientos superovulatorios que la Holstein [8] y la Hereford ms que la Aberdeen
Angus.
Categora. Las novillas responden mejor a los tratamientos superovulatorios que las vacas
[9] [10] y [11].
Edad. La respuesta ovrica declina al aumentar la edad de los animales. Lerner et al. [12]
obtuvieron un total de embriones, un nmero de ellos trans-feribles e ndices de fertilidad
significativamente menores al avanzar la edad de los animales, al igual Caral y Bernal del
Sol [13] encontraron un efecto altamente significativo de la edad sobre los porcentajes de
fertilizacin de embriones.
Estacin. En nuestras condiciones climticas los mejores resultados en los tratamientos de
superovulacin en el ganado lechero se obtienen en la poca de sequa (octubre-marzo) que
en Cuba coincide con los meses ms frescos del ao [14]. En Israel se han obtenido
respuestas superovulatorias pobres en los meses ms clidos [15].
Nivel nutricional. El estado nutricional en que se encuentran los animales al someterlos a
un tratamiento superovulatorio es decisivo para la respuesta, siendo aconsejable que el
tratamiento sea precedido de un incremento de los niveles energticos de la racin, para
garantizar una respuesta de calidad, pues est bien demostrado que una alimentacin
deficiente atenta contra la respuesta ovrica [13]. El estado de nutricin es uno de los
factores ms importantes para seleccionar una buena donante, ya que el potencial gentico
no puede manifestarse si no existe un nivel nutricional aceptable [16].
Factores inmunolgicos. La creacin de resistencia a los tratamientos con PMSG, debida
al desarrollo de antigonadotropinas [7] [17] y -[18], disminuye la respuesta de los animales
a los tratamientos repetidos con PMSG, por lo que limita su utilizacin como donantes. La
sobrestimulacin da por resultado el fracaso en recuperar embriones de buena calidad y
puede retardar el retorno al ciclo normal del estro de la donante, En los ltimos aos se han
obtenido aumentos de la respuesta superovulatoria a los tratamientos con PMSG en la
vaca, con la aplicacin de suero anti-PMSG [13][19][25][21]y[22].
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
Factores individuales. En el seno de una misma raza, parece que existen diferencias
genticas a la respuesta superovulatoria [23]. Mariana et al. [24] mostraron diferencias de
respuesta a la PMSG en medios hermanos de la raza Charolaise y plantearon que esas
diferencias pueden estar relacionadas con la tasa natural individual ,de folculos.
Feedback negativo de progeslerona sobre las gonadotropinas. Savio et al. [25]
observaron que la tasa de crecimiento fue mayor para el primero y el tercero de los
folculos dominantes, lo que implicara que durante la fase luteal media la velocidad de
crecimiento disminuya probablemente debido al feedback negativo de la progesterona
sobre las gonadotropinas. Este momento quizs sea el de mayor dificultad para estimular el
crecimiento folicular; lo que tendra implicaciones en las respuestas a los tratamientos
superovulatorios iniciados en dicho perodo. Adems de los factores enumerados. el nivel
de respuesta est influido por el nmero y el tamao de los folculos en el momento de la
estimulacin: la presencia de uno o dos grandes folculos (en la vaca, 10 mm de dimetro,
oveja y cabra 6 mm) es determinante. Un tratamiento continuo durante 2 o 3 semanas con
un anlogo de la GnRH, es capaz de limpiar el ovario de grandes folculos. Un
tratamiento superovulatorio en este momento estimular todos los folculos pequeos (3-5
mm) al incrementar las ovulaciones y disminuir la variabilidad en la respuesta [1].
TRATAMIENTOS SUPEROVULATORIOS
Originalmente para obtener la superovulacin en las vacas, se utiliz la gonadotropina
PMSG en dosis de 1 500-3 000 UI, Con posterioridad se ha utilizado anti-PMSG
monoclonal en los tratamientos con PMSG.
Actualmente se obtienen mejores resultados con hormona FSH de origen porcino (FSHp).
Algunas vacas requieren aadirle a este tratamiento 20 % de LH para lograr un resultado
satisfactorio en la superovulacin, y otras solo se pueden superovular con la aplicacin de
PMSG [6].
La dosis de FSH puede ser de 5 mg dos veces al da durante 4 o 5 das o una dosis
decreciente comenzando por dos aplicaciones de 5 mg cada una el primer da; dos
aplicaciones de 4 mg el segundo da, de 3 mg el tercero y de 2 mg el cuarto da (Tabla 6.1).
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
TABLA 6.1 TRATAMIENTOS PARA SUPEROVULACIN EN VACAS [6]
Da
10
11
12
13
14
Momento Tratamiento 1 Tratamiento 2
Tratamiento 3
a.m.
2500 UI PMSG 5 mg FSH
5 mg FSH
p.m.
5 mg FSH
5 mg FSH
a.m.
4 mg FSH
5 mg FSH
p.m.
4 mg FSH
5 mg FSH Receptora PGF 2a
a.m.
3 mg FSH
5 mg FSH
p.m.
3 mg FSH
5 mg FSH Donante PGF 2a
a.m.
2 mg FSH
5 mg FSH
p.m.
2 mg FSH
5 mg FSH
a.m.
p.m.
IA
IA
IA
a.m.
IA
IA
IA
p.m.
Celo = Da 0
IA
IA
IA
15
Recientemente se ha puesto de manifiesto una FSH de origen ovino (FSHo) con bajos
niveles de LH que es ahora preferida [26] por superar los resultados obtenidos con la FSHp
(Tabla 6.2).
La aplicacin del FSH se puede simplificar con el uso de una microbomba osmtica, que
se implanta subcutneamente y funciona de manera ininterrumpida da y noche.
TABLA
6.2
RESULTADOS
OBTENIDOS
EN
TRATAMIENTOS
DE
SUPEROVULACIN CON PMSG, FSHp Y CON FSHo EN VACAS SIBONEY DE
CUBA
Tratamiento
Respuesta (%)
Embriones
PMSG
22
81.8
Colectados (x)
4.1
Transferidos (x)
3.1
FSHp
18
77
8.2
4.2
FSHo
16
100
8.5
6.7
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
La bomba consiste en un pequeo depsito de material flexible e impermeable, rodeado
por un agente osmtico y todo est rodeado por una membrana semipermeable. Este genera
una presin hidrosttica en la pared del depsito y lo va comprimiendo gradualmente,
produciendo un flujo constante de su contenido hacia afuera, solo sale de la bomba el
material incorporado en el depsito, el agente osmtico no puede introducirse en el
depsito ni salir a travs de la membrana semipermeable. Esta microbomba osmtica ha
sido utilizada para tratamientos de superovulacin en vacas [28] y en venados [26] pero su
principal uso debe ser en trabajos de investigacin.
Tanto la PMSG como la FSH pueden ser administradas por va intramuscular o subcutnea.
Las donantes pueden ser sometidas a tratamiento de superovulacin cada 50 o 60 das,
indefinidamente. No obstante, a menudo a estas se les permite realizar un parto para que
pierdan un poco de grasa, la que les puede resultar perjudicial.
Antes de iniciar un tratamiento superovulatorio, la donante debe ser diagnosticada para
comprobar la presencia de un cuerpo lteo en un ovario, as como que no presente
anomalas en su tracto genital. Las receptoras tienen que ser diagnosticadas con los mismos
objetivos.
Sincronizacin del celo de la donante y de la receptora
La supervivencia del embrin transferido depende de la sincronizacin del celo de la
receptora con la donante. Esta sincronizacin es necesaria, para asegurar al embrin las
mismas condiciones en el nuevo tero en que continuar su desarrollo.
Algunos autores sealan los mejores resultados cuando la sincronizacin es exacta [29]
[30] o las receptoras presentan el celo poco antes que las donantes [3][6]. Pero otros han
obtenido resultados ligeramente mejores cuando las receptoras lo presentan un da despus
de las donantes (+1 da) [31] [32] [33] y [34]. Esto puede ser debido a que el embrin sufre
un retardo en su desarrollo por estrs durante el proceso de transferencia.
El mtodo ms efectivo de sincronizacin de celo en la actualidad es la aplicacin de
prostaglandina F2a. El celo se espera alrededor de 48 h despus de su aplicacin.
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
Fierra que presenta celo la donante (da 0) ya sea este natural o inducido, se aplica una
dosis de prostaglandina F2a a cada una de las receptoras, las que deben tener un cuerpo
lteo funcional. El da 10 se aplica una dosis de PMSG o en su lugar de FSH durante
cuatro das. El da 11 en las receptoras y el 12 en la donante se aplica una segunda dosis de
prostaglandina F2a. El da 14 en la tarde y el 15 en la maana y en la tarde se insemina a la
donante. Entre los das 21 y 23 se recolectan los embriones y se trasplantan a las receptoras
(Figura 6.1).
Las vacas donantes que se demoran 1 o 2 das ms de lo esperado en presentar celo, por lo
general, tienen pobres resultados. En ocasiones el celo se puede adelantar de 12-24 h. Es
usual que estas vacas tengan una buena respuesta superovulatoria y embriones de buena
calidad.
Cuando una vaca donante presenta celo durante su tratamiento superovulatorio, se debe
suspender este tratamiento y comenzar su inseminacin. Si se dispone de un crecido
nmero de receptoras, se puede prescindir de sincronizar su celo y utilizar a las que
presentan celo el da deseado.
FIGURA 6.1 Esquema de superovulacin y sincronizacin donante receptora.
Inseminacin de la donante
Las vacas sometidas a tratamiento de superovulacin, comienzan a ovular de 8-10 h
despus de concluido el celo, pero las ovulaciones se .extienden sobre 24 h, por lo que las
vacas superovuladas se inseminan tres o cuatro veces, con intervalos de 12 h entre una y
otra inseminacin. La primera inseminacin se hace de 8-12 h despus de comenzado el
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
celo. Se pueden utilizar dos dosis de semen en cada inseminacin o mejor una dosis con
mayor concentracin.
Se deben tomar todas las medidas para aumentar el nmero de embriones recolectados.
Entre ellas tenemos:
1. Emplear toros de buena fertilidad.
2. Utilizar si es posible semen lquido o monta natural.
3. Evaluar el semen antes de utilizarlo, prestando atencin a su morfologa y al porcentaje
de acrosomas intactos, as como al empleo de una buena e higinica tcnica de
inseminacin [6].
Recoleccin de embriones
El primer mtodo empleado en la recoleccin de embriones en la vaca, fue el quirrgico
(Figura 6.2), con el que se recolectaba un porcentaje alto de embriones, pero era evidente
la necesidad de modificar el procedimiento de recoleccin, por el peligro que representaba
para las donantes valiosas, los inconvenientes consecutivos a las laparatomas, en especial
las adherencias que se presentaban y que con frecuencia comprometan su fertilidad
posterior. Adems su utilizacin estaba por ello limitada a tres veces como mximo.
FIGURA 6.2 Fundamentos de la recoleccin quirrgica [4]
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
n la actualidad se utiliza el mtodo no quirrgico (Figura 6.3), que posibilita la
repetibilidad del proceso de recoleccin y la integridad de las hembras donantes; se puede
realizar en la propia instalacin ganadera donde estas se encuentran, con la limitante que
solo se puede recoger el embrin cuando se halla en el tero y que en algunas hembras no
es posible penetrar con el catter a travs del cuello, en el diestro. Los embriones son
colectados empleando este mtodo, del quinto al sptimo da, perodo en que el embrin ha
llegado al tero y an posee la zona pelcida, lo que facilita su recoleccin.
En ovejas y cabras, la recuperacin quirrgica produce adherencias que limitan la
posibilidad de repetidos lavados. Con un acceso laparoscpico las adherencias son
limitadas y la recuperacin puede ser repetida ms de siete veces en el mismo animal [1].
FIGURA 6.3 Fundamentos de la recoleccin no quirrgica [4]
FORMAS DE RECOLECCIN
La recoleccin de embriones se puede realizar en circuito cerrado o abierto. En circuito
cerrado se puede practicar con catteres de tres vas que permiten un flujo continuo, o con
catteres de dos vas que requieren un flujo discontinuo; en circuito abierto se practica, de
forma discontinua, utilizando catteres de dos vas.
Recoleccin en circuito cerrado
Cuando se utilizan catteres de tres vas se destina una a inyectar el medio del lavado en
forma continua, otra a llenar el baln con aire o con medio y una tercera va para la salida
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
del medio (Figura 6.4). Colocando el frasco con el medio aproximadamente 1 m por
encima del frasco colector se puede inyectar el lquido por gravedad.
FIGURA 6.4. Esquema de circuito cerrado con flujo continuo [35]
Con el catter de dos vas es necesario buscar una vlvula a la salida del medio que permita
extraer con una jeringuilla volmenes de 30-50 mL e inyectarlos en direccin del cuerno.
Cerrando posteriormente la vlvula el lquido que regresa del cuerno puede ser desviado en
direccin del frasco colector mediante una unin en Y (Figura 6.5).
FIGURA 6.6 Esquema de circuito cerrado con flujo discontinuo [35]
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
Recoleccin en circuito abierto
Para la recoleccin en circuito abierto se utilizan catteres de dos vas, una destinada a la
inyeccin de aire y la otra a la inyeccin y recoleccin del medio, con una jeringuilla de 50
mL. Durante el tiempo que se emplea en recargar con medio la jeringuilla despus de cada
lavado, el catter debe permanecer cerrado con una pinza para evitar que se pierda el
lquido que puede quedar en el cuerno. Se deben practicar alrededor de 15 o 20 lavados.
Esta forma de recoleccin no requiere del uso de vlvulas ni uniones en el catter (Figura
6.6).
FIGURA 6.6 Esquema de circuito abierto con flujo discontinuo. Inyeccin y recoleccin con jeringuilla [35]
Ventajas y desventajas de estas formas de practicar el lavado
Aunque el circuito cerrado se supone que tenga menos peligro de contaminacin, tiene la
posibilidad de penetracin de grmenes por una desinfeccin insuficiente, de un
instrumental ms complejo o de contaminacin de productos qumicos por fallos en el
proceso de desinfeccin.
La forma de recoleccin en circuito abierto de forma discontinua tiene la ventaja de que se
emplean medios ms simples con menos riesgos de contaminacin, pero solo se puede
utilizar en animales dciles y en condiciones adecuadas [35].
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
CATTERES
Los lavados uterinos para la recoleccin no quirrgica de embriones originalmente se
hacan con catteres Foley de dos o de tres vas. Estos presentan serias dificultades tales
como el resultar cortos para vacas grandes, ser de goma blanda, lo que dificulta su
introduccin a travs del cuello uterino, as como poca distancia desde el baln hasta el
extremo de salida del medio utilizado para el lavado. No obstante, estas dificultades, su
fcil obtencin y precio barato, motiva su empleo por muchos grupos que trabajan en
transferencia de embriones.
Existe otro catter, el de Rusch de dos vas modelo NEUSTAD/AISCH de 67 cm de largo y
cuatro orificios en la punta, el que es endurecido con un estilete de acero inoxidable para
facilitar su introduccin a travs del cuello uterino. Resulta suficientemente largo para su
uso en vacas grandes y pasa fcilmente al lumen uterino, lo que le ha valido su aceptacin
por muchos grupos.
Newcombk [29] desarroll un catter largo para lavados uterinos, con 42,5 cm desde el
baln hasta el extremo donde se encuentra el orificio de salida. El lavado es recuperado a
travs de un 'orificio situado inmediatamente por delante del baln. Segn este autor con;
el referido catter se pueden recolectar ms embriones que con los catteres
convencionales.
Los catteres deben ser esterilizados con xido de etileno o una solucin esterilizante dbil.
Cuando se utiliza el xido de etileno es indispensable que el catter est seco y despus
aerearlo durante una semana, porque puede matar los embriones. Similarmente las
soluciones esterilizantes dbiles que se utilicen son embriotxicas, requiriendo que el
catter sea lavado en solucin salina estril, despus de su aplicacin.
MEDIO
El medio usualmente empleado para la recoleccin de embriones es el PBS, que es una
solucin salina de fosfato buferada, con un 2 % de suero fetal bovino inactivado por calor
y debe ser atemperado a la temperatura corporal cuando se va a utilizar. El medio se
conserva refrigerado en frascos de 500 mL listo para ser usado. El suero de temeros recin
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
nacidos, libre de inmunoglobulinas, es similar al suero fetal y puede ser usado para
sustituir a este ltimo, con ventaja desde el punto de vista econmico.
Bsqueda de los embriones
Independientemente del mtodo de recoleccin que se utilice, es necesario dejar reposar al
medio en un embudo separador, durante 35 min como mnimo, para que los embriones
vayan al fondo. La extraccin de estos del embudo separador se puede realizar, mediante
sifonaje con un tubo de silicn estril, del medio depositado en el embudo separador, hasta
dejar los ltimos 50 mL en el fondo -teniendo mucho cuidado para no agitar el medio, lo
que puede causar prdida de embriones. El remanente que se deja en el embudo separador
se mueve suavemente y se aspira mediante una pipeta de infusin uterina y se deposita en
una placa de Petri (100 x 15 mm). Otra alternativa es aspirar 35 mL del fondo del embudo
separador despus de que el medio est asentado, y repetir la operacin dentro de 10 o 15
min con lo que se obtienen buenos resultados.
La placa con el medio se debe observar en un microscopio esteroscpico en cuya base se
puede colocar una rejilla con cuadrculas del tamao de un campo de microscopio. Si se
utiliza una placa de material plstico se raya en el fondo para hacerle las cuadrculas.
Los campos se deben observar manteniendo un orden, el que se puede establecer a
voluntad del operador, con un aumento de 10 dimetros, Cuando se termina la observacin
se debe mover la placa para desituar los embriones que estn en los bordes de la placa y
buscar de nuevo. Cada placa debe ser observada al menos dos veces despus de que el
ltimo embrin fue encontrado, repitiendo el movimiento para cambiar de posicin a los
embriones en cada caso [6].
Los embriones a medida que son localizados se extraen con un catter plstico calibre 20
unido a una jeringuilla de 1 mL y depositados en una pequea placa de Petri (35 x 15 mm)
que contiene medio de cultivo. Este es un medio semejante al del lavado uterino pero con
la adicin de un 10-20 % de suero fetal bovino. Cuando termina la bsqueda los embriones
son evaluados con un aumento de 50-100 dimetros.
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
CARACTERSTICAS DE LOS EMBRIONES
El tamao de los embriones durante los 8 o;9 primeros das permanece virtualmente sin
cambios (150-190 ) incluyendo la zona pelcida (12-15 ), entonces salen de dicha zona
(Figura 6.7). Es fcil distinguir los huevos que no fueron fertilizados de los fertilizados y
los estadios por los que estos pasan.
FIGURA 6.7 Embrin bovino de 14 das fuera de la zona pelcida.
Ovocitos no fertilizados. La presencia en el medio utilizado en el lavado uterino, de
ovocitos con una sola clula, indica que se trata de ovocitos que no fueron fertilizados
(Figura 6.8). Estos ovocitos no tienen un contorno celular definido. Presentan material
celular disperso en el espacio perivitelino, con frecuencia presentan detritus celulares
adheridos a la zona pelcida, especialmente en los lavados infectados.
FIGURA 6.8 Ovocito de 10 das no fertilizado
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
Ovocitos fertilizados. Los ovocitos fertilizados (Figura 6,9) [36] pasan por los siguientes
estadios en su desarrollo:
Estadio precoz de la divisin celular. Se desarrolla durante su trnsito por el oviducto en el
que se produce su divisin en dos, cuatro, ocho y dieciseis clulas.
Mrula. Presenta una masa de al menos 16 blastmeros difciles de diferenciar unos de
otros, que ocupa la mayor parte del espacio perivitelino.
Mrula compacta. Las blastmeras se unen formando una masa compacta que ocupa ms
del 60 o 70 % del espacio perivitelino.
Blastocisto joven. Comienza a formarse el blastocele, pequea cavidad llena de lquido. El
embrin ocupa del 70-80 % del espacio perivitelino.
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
FIGURA 6.9 Estadios del desarrollo del ovocito fertilizado [36]
Blastocisto. El blastocisto est bien formado. Las clulas del trofoblasto estn bien
diferenciadas de la masa compacta de clulas. Es espacio periviteiino prcticamente
desaparece. Las clulas del botn embrionario son visibles.
Blastocisto bien formado. El dimetro del embrin aumenta dramticamente. La zona
pelcida se adelgaza ms o menos 1/3 de su grosor original.
Evaluacin de los embriones
En el lavado que se realiza en el tero para recolectar los embriones, se pueden encontrar
huevos que no fueron fertilizados y embriones en distintos estadios de su desarrollo, que
pueden ir desde mrulas hasta blastocistos que salen de la zona pelcida. Es probable que
esta diferencia en los estadios de desarrollo en que se hallen los embriones, obedezca a
ovulaciones que se produjeron en distintos momentos tras el tratamiento superovulatorio.
En el lavado igualmente se pueden encontrar embriones de excelente calidad, esfricos,
simtricos con clulas de tamao uniforme, junto a embriones que no muestran un
desarrollo completamente normal, que puede comprometer su posterior gestacin en
receptoras. No obstante, algunos embriones de calidad pobre pueden sobrevivir en
receptoras bien sincronizadas con la donante.
CLASIFICACIN DE LOS EMBRIONES
Los embriones se pueden clasificar [6] en:
Excelentes. Un embrin ideal, esfrico, simtrico y con clulas de tamao, color y textura
uniforme (Figura 6.10).
Buenos. Presentan pequeas imperfecciones, como unas pocas blastmeras de forma
irregular y algunas vesculas desprendidas.
Regulares. Problemas ms definidos que incluyen blastmeras desprendidas, vesculas y
algunas clulas degeneradas.
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
Malos. Embriones que presentan problemas severos, como numerosas blastmeras
desprendidas, clulas degeneradas, clulas de variables tamaos, numerosas vesculas, pero
una aparente masa de embrin viable. Estos embriones generalmente no son de calidad
para transferirlos.
FIGURA 6.10 Embriones de excelente y buena calidad
Implantacin de embriones
La implantacin de embriones o embrionizacin, al igual que la recoleccin de embriones,
se puede realizar por mtodos quirrgicos o por mtodos no quirrgicos.
MTODO QUIRRGICO
Existen dos tcnicas fundamentales para la implantacin quirrgica, una por la lnea media
y otra por el flanco.
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
Implantacin por la lnea media
La receptora es sometida a anestesia general o local y colocada en decbito lateral. Se
prepara el campo operatorio en la regin ventral, delante de la glndula mamaria, se
penetra en la cavidad abdominal por incisin en la lnea media. Se localiza el tero, se
confirma la presencia del cuerpo lteo en uno de los ovarios y se hace una pequea puntura
en el cuerno correspondiente al ovario con el cuerpo lteo y se introduce una pipeta que
contiene el embrin y se deposita este en el lumen uterino. Por ltimo, se sutura la herida.
Es conveniente ayuno de comida de 24-48 h y de agua de 12-24 h antes de esta operacin.
Implantacin por el flanco
La implantacin del embrin por el flanco, tiene la ventaja de poderse realizar con el
animal de pie. Se diagnostica la receptora por va rectal para determinar en qu ovario se
encuentra el cuerpo lteo y operar por el flanco ms prximo a este. Despus de incidir los
tejidos se aproxima con cuidado el cuerno ipsilateral al cuerpo lteo lo ms cerca posible
de la incisin y se procede a depositar el embrin en el lumen uterino en la misma forma
descrita en la implantacin por la lnea media, procediendo posteriormente a la sutura de
los tejidos.
Mtodo no quirrgico
Distintos instrumentos y tcnicas "se 'han desarrollado para realizar la implantacin de
embriones no quirrgica, los ms usados son los modelos japoneses y franceses en especial
el de CASSOU.
El embrin es situado en la pajuela o en el tubo de polietileno que se vaya a introducir en
el tero, entre dos burbujas de aire para protegerlo de los movimientos.
La receptora es explorada para determinar la presencia y localizacin de un cuerpo lteo
apropiado. Se aplica anestesia epidural baja para evitar las contracciones del recto, se
limpia bien y se seca la vulva y sus alrededores. Se introduce el instrumento que se vaya a
utilizar y se deposita el embrin dentro del cuerno ipsilateral al cuerpo lteo.
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
Algunos depositan el embrin por delante de la bifurcacin uterina externa y otros tratan
de hacerlo ms craneal. Es preferible no depositarlo tan cranealmente, antes que realizar
una dificultosa y prolongada deposicin ms craneal [3].
La tcnica no quirrgica tiene la ventaja sobre la quirrgica de realizarse en menos tiempo,
no requerir facilidades especiales de instalacin, y tener un menor costo. Las desventajas
son la prolongada preparacin tcnica del personal que la va a realizar, para adquirir la
habilidad necesaria y el hecho de que algunas receptoras presentan gran dificultad para
penetrar a travs del cuello uterino [3]. Esta dificultad es posible prevenirla con la
aplicacin unas horas antes de la implantacin de 10 mL de una solucin al 2 % de
lidocana en el crvix de la receptora.
En general la va cervical es la va de eleccin en la vaca, por su comodidad. Esta est
ampliamente generalizada y los niveles de fertilidad obtenidos son de 50 % como
promedio. La transferencia quirrgica en los cuernos uterinos o en el oviducto pueden
aumentar la fertilidad en un 10 o un 15 % y es utilizada excepcionalmente para los
embriones de alto valor gentico o de alto precio.
En la oveja y en la cabra la va cervical no es utilizada en razn de sus particularidades
anatmicas. La transferencia de embriones es realizada en estas especies por va quirrgica
clsica o cada vez ms por endoscopia [37].
Aspectos epizootiolgicos de la transferencia de embriones
Debido a la difusin de la transferencia de embriones en el mundo y el intercambio
internacional de los embriones, se hace necesario considerar las posibilidades de
transmisin de enfermedades por los embriones. Segn un estudio de la Sociedad
Internacional de Transferencia de Embriones, la capacidad de transmitir enfermedades
infecciosas de los embriones es considerablemente menor que la posibilidad de
transmitirlas por el semen o por los animales vivos [25].
Es indispensable respetar las reglas sanitarias del lavado de embriones: utilizacin de
embriones con la zona pelcida intacta y libre de restos celulares, aplicacin de
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
antibiticos al medio de coleccin de lavados y al transporte de embriones y trasplante
uterino en condiciones sanitarias.
Un nmero suficiente de transferencias efectuadas de embriones provenientes de donantes
infectadas con el virus de la leucosis bovina, de taringotraqueitis infecciosa bovina (IBR),
de la enfermedad de las mucosas, as como de donantes porcinas infectadas con el virus de
la enfermedad de Aujeski puede afirmar que estos virus no son transmitidos por los
embriones a condicin de que sean manipulados correctamente. Se debe insistir en que la
posibilidad de prevenir la transmisin de enfermedades por los embriones depende del
cumplimiento de las medidas establecidas para ello y que si estas son efectuadas
impropiamente pueden contribuir por el contrario a introducir agentes infecciosos en los
rebaos [37].
Conservacin de embriones
Si el tiempo que transcurre entre la recoleccin y la transferencia del embrin, se logra
reducir a 1 o 2 h, optimizando las condiciones de rutina, no es necesario el empleo de
medios especiales de conservacin, para intervalos mayores, 6 -12 h, se requiere utilizar
medios de cultivo y los resultados son variablemente menores.
Cuando se necesita consumir de 24-36 h, se puede conservar de 0-4 C, con lo que se
obtienen porcentajes normales de gestacin. El estadio del ciclo de la receptora puede ser
el mismo que el de la donante el da de la recoleccin.
Sobre 48 h de conservacin afecta los resultados.
La conservacin por perodos largos requiere la congelacin del embrin con nitrgeno
lquido (-196 C).
CONGELACIN DE EMBRIONES
Uno de los mayores problemas en la transferencia de embriones es la correspondencia
entre embriones disponibles y receptoras sincronizadas. En el momento de realizar las
transferencias pueden sobrar embriones por haberse recolectado ms de los calculados o
sobrar receptoras sincronizadas por haberse colectado menos embriones. Ambas
dificultades pueden ser resueltas con la utilizacin de la congelacin de embriones. Otras
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
ventajas de la congelacin de embriones son la de poder conservarlos durante meses o aos
sin que pierdan su viabilidad, su facilidad de transportacin, poco espacio de
almacenamiento y el de poder obtener cras despus de muertos sus progenitores [38].
Adems facilita el poder aprovechar la ventaja que ofrece la transferencia de embriones de
obtener cras sanas procedentes de rebaos o de receptoras afectadas de enfermedades
infecciosas y posibilita el comercio internacional de embriones.
En el bovino, los mejores resultados de supervivencia a la congelacin se obtienen con
blastocistos precoces (7-8 das). En la oveja, la etapa del desarrollo embrionario no es un
factor tan importante para la supervivencia posterior a la congelacin, lo mismo sucede con
conejos y ratones.
La congelacin requiere la utilizacin de un crioprotector, glicol para los embriones del
bovino, etilenglicol para los ovinos, uno u otro para los de los caprinos y propanodiol para
los humanos y los del conejo.
El cambio de fase lquida a slida debe ser cuidadosamente controlado. El descenso de
temperatura se hace rpidamente de + 20 a - 7 C a razn de 5 C/min. De -30 a -35 C el
descenso debe ser excesivamente lento a razn de 0,3 C/min, para que la deshidrataron del
embrin sea lo suficientemente alta para disminuir la cristalizacin en el interior de las
blastmeras al sumergirlo en el nitrgeno lquido, la inmersin debe ser instantnea (Figura
6.11) [37].
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
FIGURA 6.11 Programa de congelacin utilizado para los embriones bovinos [36]
DESCONGELACIN DE EMBRIONES
La descongelacin debe ser preferiblemente rpida, en agua a 20 o 37 C. El agente
crioprotector debe ser removido de las clulas, esto usualmente es logrado por 4-6 lavados
sucesivos en soluciones de concentracin decreciente del crioprotector. Durante esta
manipulacin, los embriones son examinados y solo aquellos que han sobrevivido son
transferidos.
Esta tcnica ha sido usada por casi todos los equipos de transferencia de embriones del
mundo. Permite una sobrevivencia de un 85-95 % de embriones, dependiendo del curso y
de cmo fue hecha la seleccin precongelacin. Los resultados no difieren de los obtenidos
con embriones frescos (50-60 %).
Una tcnica mucho ms simple se logra con el uso de molculas como sacarosa que por no
penetrar en las clulas permite la eliminacin del crioprotector en un solo paso. En efecto
la sacarosa no penetra las clulas y aumenta la presin osmtica del medio extracelular del
embrin y provoca la difusin pasiva del crioprotector en el interior de las clulas
embrionarias y el blastocele [39].
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
Adems, si la pajuela en que fue congelado el embrin contiene tambin un volumen de
solucin de sacarosa, la transferencia puede ser hecha de manera directa en la granja, sin
examen previo, tan simplemente como realizar la inseminacin artificial [39]. La misma
propiedad de la sacarosa puede ser usada para simplificar el proceso de congelacin.
Esquemas de seleccin gentica MOET
La inseminacin artificial ha hecho posible las pruebas de progenie para valorar la calidad
gentica del macho, de gran importancia en la mejora ganadera. Mediante el trasplante de
embriones y las ovulaciones mltiples (MOET) se puede poner a prueba la calidad
gentica sobre la base de gametos femeninos y valorar del mismo modo las hembras,
pudiendo disponer igualmente de hembras probadas o mejoradoras. Los resultados de estos
avances sern capaces de acelerar de manera insospechada la mejora ganadera y los
rendimientos cuantitativos y cualitativos de las producciones pecuarias.
El progreso gentico es ms rpido cuando se utilizan esquemas de seleccin asociados a la
superovulacin y el trasplante de embriones (MOET), que por medio de los procesos
convencionales de chequeo de la descendencia. El intervalo de generacin pasa de ms de
seis aos a cuatro o cinco y hasta dos segn los diferentes esquemas MOET (Tabla 6.3).
TABLA 6.3 PROGRESOS GENTICOS ANUALES EN PRODUCCIN LECHERA
BOVINA POSIBLES CON DIFERENTES ESQUEMAS DE SELECCIN
Sistemas de esquemas de seleccin
Progreso cientfico
Intervalo de
(unidades SD x 100)
100
generaciones (anuales)
6.3
126
5.6
MOET de hembras adultas
130 a 145 b
5.1
MOET de hembras jvenes
141 a, 154 b
4.2
MOET de adultos de ambos sexos
118 a, 158 b
3.7
MOET de jvenes de ambos sexos
178 a, 255 b
1.8
Prueba
tradicional
sobre
la
descendencia
Prueba eficaz sobre la descendencia
Prueba de descendencia de machos con:
Esquemas de grupos bovinos base:
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
(a) Esquema MOET normal: 4 machos y 4 hembras son seleccionados, con 8 donantes ce
ovocitos por macho.
(b) Esquema MOET intensificado: 8 machos y 8 hembras son seleccionados, con 16
donantes de ovocitos por macho.
En los grupos de base de seleccin pueden estar reunidos los mejores animales
seleccionados a partir de la poblacin. Esto quiere decir que las bases parten de un
potencial gentico netamente ms elevado que el resto de la poblacin. Estos animales
pueden ser mejor controlados bajo criterios mltiples de seleccin [37].
La desventaja de esquemas genticos de unidades bases est dada por lo pequeo de los
grupos formados, sobre todo, en comparacin nacional y la rotacin ms rpida de las
generaciones, que elevan la tasa de consaguinidad cuando los esquemas MOET se utilizan
a partir de animales jvenes. No obstante si existen muchas bases de seleccin con
intercambio entre ellas, los problemas de consanguinidad pueden ser sustancialmente
disminuidos [37].
Fecundacin in vitro
La primera fertilizacin in vitro (FIV) confirmada fue obtenida en el conejo en 1954, al
fecundar a los ovocitos fertilizados, con espermatozoides capacitados en el tero, y cinco
aos ms tarde se confirm la normalidad biolgica de FIV al obtener un nacimiento.
En el humano se logr la primera fertilizacin en 1969 y el primer nio probeta naci en
1978, y en bovinos la fertilizacin del ovocito en 1980 y el nacimiento en 1982 (Tabla 6.4).
TABLA 6.4 HISTORIA DE LA FIV EN MAMFEROS [40]
Primeras fecundaciones in vitro
1954 Conejo
1963 Hmster
1968 Ratn
1969 Humano
1980 Bovino
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
1989 Equino
Primeros nacimientos
1959 Conejo
1978 Humano
1982 Bovino
1985 Caprino
1986 Ovino
1990 Equino
Cuba fue el primer pas en Amrica Latina y el duodcimo pas en el mundo en obtener un
nacimiento en bovinos, en agosto de 1992.
La manipulacin in vitro de los gametos tiene como base el fundamento natural, que el
ovocito es ovulado antes de su evolucin completa en el folculo de De Graaf -metafase de
la segunda meiosis y los espermatozoides tampoco terminan el ciclo de madurez hasta
abandonar el genital masculino [41].
Para la fecundacin in vitro, no solo se necesita la capacitacin espermtica, adems, se
requiere la maduracin completa del ovocito, tanto nuclear como citoplasmtica. Las
modificaciones que sufre la zona pelcida durante el perodo preovulatorio tambin debe
influir en el proceso de fertilizacin [40].
En los roedores la capacitacin se obtiene por una corta incubacin de los espermatozoides
en un medio relativamente simple, pero en e bovino requiere la adicin de heparina al
medio bsico. Si el espermatozoide no est completamente capacitado cuando se aade a
los ovocitos, la fertilizacin se demora muchas horas, lo que da lugar al envejecimiento del
ovocito antes de la fertilizacin. Los que envejecen pueden sufrir anomalas del desarrollo.
Existen grandes variaciones en los resultados obtenidos con la FIV cuando se utilizan
eyaculados de diferentes machos. Estas variaciones se pueden reducir en los bovinos al
ajustar las concentraciones de heparina de cada eyaculado. Algunas condiciones como la
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
temperatura tambin son crticas, se requiere una temperatura de 39 C en bovinos, ovinos
y suidos (Tabla 6.5).
TABLA 6.5 INFLUENCIA DE LA TEMPERATURA SOBRE EL PORCENTAJE DE
FERTILIZACIN [40]
Especie
Cerda
Temperatura (C)
37
Fertilizacin (%)
1
39
89
37
39
53
33
39
80
37
13
Vaca
Oveja
Coneja
38.5
86
La demora en obtener resultados con la FIV en las grandes especies de animales
domsticos, se explica por la necesidad de utilizar eyaculados en lugar de espermatozoides
obtenidos del epiddimo, que tienen una mayor habilidad para capacitarse, pero usualmente
requiere el sacrificio del animal para obtenerlos. Adems, estas especies necesitan mayor
tiempo para la capacitacin del esperma que los roedores y el hombre, por lo que es
indispensable tener condiciones del cultivo que mantengan por muchas horas la
supervivencia y la motilidad de los espermatozoides [40].
La posibilidad de observar los dos proncleos, macho y hembra, durante la FIV, permite
aplicar las manipulaciones fundamentales del genoma utilizadas en el estudio de
fenmenos biolgicos fundamentales, tales como el clonaje, la transgnesis, la
partenognesis, etctera (Figura 6.12) [37].
Respecto a la produccin in vitro de embriones, la tecnologa animal fue por detrs de los
avances de la medicina humana; sin embargo, en la ltima dcada se ha invertido el signo
del progreso de manera muy destacada. El planteamiento se ha hecho, sobre la base de la
obtencin de ovocitos de vacas sacrificadas, para luego ser madurados, fecundados y
conservados hasta su utilizacin en trasplantes [41].
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
RECOLECCIN DE OVOCITOS DE HEMBRAS SACRIFICADAS
Los ovarios de vacas sacrificadas se trasladan al laboratorio en una solucin PBS a
temperatura de 24-25 C, lo que permite conservarlos durante 11 h sin que sus ovocitos
pierdan su capacidad de ser fecundados ni del posterior desarrollo de los embriones.
Despus de lavar los ovarios tres veces con el medio de PBS se procede a obtener los
ovocitos de todos los folculos terciarios que poseen un tamao entre 2 y 6 mm de
dimetro, mediante la puncin con un aparato que opera con una presin negativa de 100
mm Hg. Si no se cuenta con dicho aparato se puede aspirar los ovocitos con una
jeringuilla. La aguja en ambos casos es de 0,9 mm de dimetro. El lquido folicular con los
ovocitos se deja sedimentar aproximadamente 15 min antes de aislar los complejos
cumulus-ovo-cito (Figura 6.13).
FIGURA 6.12 La FIV permite importantes manipulaciones del genoma [37]
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
FIGURA 6.13 Esquema de la produccin in vitro de embriones. Obtencin y maduracin in vitro de los
ovocitos [39]
Se destinan a la maduracin los ovocitos que posean un cumulas oophorus denso que cubra
la totalidad del ovocito y un citoplasma uniformemente granulado y oscuro [42]. Los
ovocitos que cuentan con un cumulas completo y compacto poseen la capacidad de
completar la maduracin In vitro.
Por animal se obtiene un promedio de 15-18 ovocitos de los cuales entre el 57 y 70 %
responden a los criterios morfolgicos para una exitosa maduracin in vitro [42].
IMPORTANCIA DE LA FECUNDACIN IN VITHO
La FIV posibilita la utilizacin de un considerable nmero de ovocitos obtenidos
posmortem, de animales de gran valor gentico, muertos por accidente o sacrificados al
llegar al final de su vida productiva.
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
Adems, permite la utilizacin de ovocitos obtenidos de ovarios de donantes vivos y
constituye un paso importante de mltiples micromanipulaciones de genoma,
Quizs la perspectiva ms interesante que ofrece en este momento la FIV est relacionada
con la posibilidad de reducir como mnimo en la proporcin de 1: 1000 los
espermatozoides necesarios para fecundar un ovocito [41]. Esto representa multiplicar
muchas veces el aprovechamiento del semen de sementales muy valiosos.
Clonaje
Consiste en la obtencin de un gran nmero de copias rigurosamente iguales de un animal
o planta de alto valor gentico. Todos los gemelos constituyen un clon, del griego KIon
(pequea rama).
Casi todas las clulas de un organismo contienen la. informacin gentica necesaria para
reproducir un organismo complejo. El principal problema es que esta se exprese. Hasta
ahora esto se haba logrado con clulas inmaduras totipotentes, ya que a medida que las
clulas se hacen diferenciadas parecen perder la habilidad de emitir instrucciones genticas
hacia otra cosa que no sea en lo que ellas se han convertido, no obstante, recientemente en
Irlanda se obtuvo el nacimiento de la oveja Dolly a partir de una clula de la mama.
Tecnologa de la transferencia nuclear
Los ovocitos obtenidos por superovulacin y maduracin in vitro son liberados de los
restos del cumulus oophorus y enucleados; el material nuclear, constituido por
cromosomas en metafase II, y el primer corpsculo polar, son succionados por medio de
una micropipeta ms fina que un cabello humano a travs de la membrana pelcida.
Los cromosomas se pueden ver por fluorescencia. Despus de enucleados los ovocitos son
tratados con citocalasine B, que aumenta de forma reversible la elasticidad de la membrana
y evita su ruptura. Las blastmeras de un embrin donante (4-64 clulas) son ahora
colocadas bajo la membrana pelcida. Por electrofusin de la membrana plasmtica, el
ncleo de la blastmera penetra en el citoplasma del ovocito receptor enucleado. La
electrofusin a la vez permite la activacin del ovocito receptor, etapa indispensable a la
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
reprogramacin del ncleo donado, que ya haba recibido la activacin de su genoma
embrionario.
En la oveja, la coneja y la cerda, los embriones activados por la electrofusin, son
trasplantados quirrgicamente al oviducto de hembras receptoras con su ciclo sincronizado.
En bovinos se han empleado como intermediarias ovejas y conejas, en cuyos oviductos
ligados se ha comenzado a desarrollar el embrin, o cilindros de agar-agar con el mismo
propsito. Despus de 5 o 6 das los embriones son trasplantados a las vacas receptoras,
son congelados con vista a su ulterior transferencia, o a su vez servir de donantes de
ncleos [37].
Repitiendo este proceso con las blastmeras de los embriones destinados a servir de
donantes, se puede obtener el nacimiento de clones de terneros (Figura 6.14).
FIGURA 6.14 Clonaje de embriones [37]
Transgnesis
Un animal transgnico es el que ha adquirido una nueva informacin gentica como
resultado de la introduccin de ADN extrao a su propio patrimonio gentico [37].
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
La transgnesis se logra al incorporar ADN exgeno al genoma de una clula totipotente,
que puede ser un ovocito fertilizado o una clula embrionaria no diferenciada.
La introduccin del gen puede ser directa, por microinyeccin del ADN dentro del
proncleo de un huevo fertilizado o por infeccin de un embrin con un retrovirus
recombinante, o puede ser indirecta, aprovechando la ventaja de clulas embrionarias
totipotentes, derivadas de blastocistos adaptados a cultivos de tejidos (Figura 6.15). Estas
lneas de clulas si son reintroducidas dentro de mrulas o blastocistos pueden colonizar el
embrin y participar en su desarrollo [43].
Cuando la integracin del ADN exgeno es por recombinacin heterloga, el resultado es
la adicin de un gen al genoma. Si la integracin es por recombinacin homologa, el
resultado es el cambio de una porcin del gen por una nueva.
Adems de ratones es posible obtener otros mamferos transgnicos. No obstante la
eficiencia contina baja, la microinyeccin del huevo solo ha sido obtenida por
recombinacin homologa [43]. La infeccin por retrovirus recombinante de casi todos los
mamferos parece posible. Pero al parecer solo los genes transferidos por recombinacin
homologa pueden asegurar las garantas funcionales y las ticas requeridas para la
produccin de los animales de granja.
Desde 1980 la tecnologa de animales transgnicos ha sido utilizada para estudios de
biologa molecular, diferenciacin celular, inmunologa, fisiologa, neurobiologa y
cancerologa.
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
FIGURA 6.15 Manipulacin gentica del embrin de ratn [43]
Recientemente se han establecido varios cientos de lneas de ratones transgnicos y unas
pocas de conejos, ovejas y cerdas, pero hoy da es necesario microinyectar alrededor de
100 huevos de ratn para obtener un ratn transgnico. Para obtener un conejo se requiere
el doble, treinta veces ms para obtener un cerdo, cien veces ms para una oveja o una
cabra y mil veces ms para lograr un bovino.
En produccin animal la transferencia de genes es solo interesante cuando la estructura
gnica es transmitida a la descendencia, de forma tal que se establezcan las lneas
transgnicas en la poblacin [44].
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
HERENCIAS BIBLIOGRFICAS
1).
CHEMINEAU, R; D. CHUPN; Y COGNIE; and J. THIMONIER: Control of
reproduction in domestic mammals. En: Reproduction in mammals and man. (Eds.)
C. Thibault; M.C. Levasseur and R.H.F. Hunter, Ellipses, Paris, 1993.
2).
MAPLETOFT, R.J.: Embrion transfer and genetic engineering. Introduction. En:
Current therapy in therogenology 2. (Ed.) D.A. Morrow., W. B. Saunders Company,
Philadelphia, London, Toronto, 1986.
3).
ROWE, R.F.: Transfer of bovine embryos. En: Current therapy in theriogenology 2.
(Ed.) D.A. Morrow, W.B. Saunders Company, Philadelphia, London, Toronto, 1986.
4).
CARAL, J.R y L. HOLY: Algunas consideraciones sobre la tcnica del trasplante de
embriones y su aplicacin en la explotacin del ganado bovino. En: Transferencia de
embriones. CIMA, La Habana, 1988.
5).
FOOTE, R.H. and H. ONUMA: Superovulation, ovum collection, culture and
transfer. A Review. J. DairyScL, 53(12): 1681-1692, 1975.
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
6).
MAPLETOFT, R.J.: Bovine embryo transfer. En: Current teherapy in
theriogenology 2.(Ed.) D.A. Morrow, W. B. Saunders Company, Philadelphia,
London, Toronto, 1986.
7).
MAULEON, R; HUY N. NGUYEN y N. CHUPN: Naissance gemellaire chez le
bovins. Bulltech. CWZ, 257, 1971.
8).
GORDON, I.: Problems and prospects in cattle egg transfer. Irish Vet J., 29:2130,1975.
9).
LINDSELL, C.E.; B.D. MURPHY and R.J. MAPLETOFT: Superovulatory and
endocrine responses in heifers treated with FSHp at different stages of the estrus
cycle. Theriogenology, 26(2): 209, 1986.
10). MARTNEZ, JOSEFA: Superovulacin en el ganado bovino. Efecto de la categora
de la donante sobre la respuesta ovrica. Trabajo de Diploma, ISCAH, La Habana,
1984.
11). WARFIELD, S.J.; G.E. JR. SEIOEL and R.P ELSDEN: A comparison of two
regimens for superovulating cows and heifers. Theriogenology, 25(1); 213,1986.
12). LERNER, S.R; W.V THAYNE; R.D, BAKER et al.: Age, dose of FSH and others
factors affecting superovulation in Holstein cows. J. of Anim. Sci., 63(1):176,1986,
13). CARAL, J.R y A. BERNAL DEL SOL: La superovulacin en el ganado bovino.
En: Transferencia de embriones. CIMA, Establecimiento Imprenta de la Empresa de
Aseguramiento y Servicios del MINAGRI, La Habana, 1988.
14). CARAL, J.R: Trasplante de embriones. Ffev. ACPA, No-2, 1984.
15). ALMEIDA, A.R: Superovulation in cattle: A combined treatmente using syncromate
B with either. PMS or FSH. Theriogenology, 27(1): 203, 1987.
16). AUGHTRY, J.D.: Nutrition of donor and recipient cows. Embryo Transfer., 2(3),
1987.
17). DELONG, K.H, and R.M. SHARPER: Evidence for inhibing like activity in bovine
follicular fluid. Nature, 263: 71-72,1976.
18). HAFEZ, E.S.E.; E. RAJAKOSKI; PB. ANDERsoNefa/.: Problems of gonadotropin
induced mltiple pregnancy in beef cattle. Amer. J. Vet. Res., 25: 1074-1079,1964.
19). BINDON, B.M. and L.R. PIPER: Pnysioiogical basis of the ovaran responso to
PMSG in cattle, sheeps and goats. (Ed.) Shelton et al-, Aust Society Reproductivo
BiologyPublication, 1982.
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
20). DIELEMAN, S.J.; M.M, BEVERS and J. TH. GIELEN: Increase of the numberof
ovulation in PMS/PG treated cows by administration of monoclonal anti PMSG
shortiy afterthe endogenous LH peak. Theriogenology, 27(1):222, 1987.
21). KIM, H.N.; R.W. RORIE; C.R.YouNcsta/.: Theuseofanti-PMSGantibodies with
PMSG for superovulating beef cattle. Theriogenology, 27(1):243, 1987.
22). MOYAERT, I.;
R.
BOUTERS;A.T.M.
SHONNERpefa/.:
The
control
of
superovulation on the bovine with a monoctonal PMSG antibody. Theriogenology,
23(1): 210,1985.
23). MAULEON, R y D. CHUPN: Reunin de la Maitrise des Cycles Sexuele chez les
Bovins. 12-13 Janvier 26-37 (Ed.) Searle Pauz, 1976.
24). MARIANA, J.C.; R GIRARD and D. CHUPN: Differences intraraces de response
ovarienne a traitment de superovulation par PMSG chez des vaches de race
Charolaise. Frecuence de naissance gemellalres. Ann. Zootech., 26(3): 345353,1977.
25). SAVIO, J.D.; L KEENANAN; M.R BOLAND and J.F. ROCHE: Pattern of growth
of dominant follicles during tree estrous cycle of heifers. J. firepot. Frtil, 83:663671, 1988.
26). FENNESSY, RF; G.W. ASHER; N.S. BEATSON et al.: Embryo transfers in deer.
Theriogenology, 41:133-138,1994.
27). BERNAL DEL SOL, A.: Informacin personal, 1997.
28). WUBISHET, A.; D.J. KESLER; C.N. GRUES et al.: Preovulatory LH profiles of
superovulated cows and progesterone concentrations at embryo recovery.
Theriogenology, 35(2): 451-457, 1991.
29). NEWCOMB, R. and L.E.A. ROWSON: Conception rate after transfer of cow eggs,
in relation to synchronization of estrus and age of eggs. J. feprod. Fertile, 43: 539,
1975.
30). ROWSON, LEA; R.A.S. LAWSON; R.M. MOR and A.A. BAKER: Eggs transfer in
the cow. Synchronization requeriments. J. Reprod. Frtil., 28: 427, 1972.
31). ALCAL, L.; R. DE ARMAS; J.R CARAL y R. SOLANO: Comportamiento de dos
tratamientos de sincronizacin astral con prostaglandina F2a en hembras receptoras.
Resmenes III Congreso Cubano de Ciencias Veterinarias. I Seminario Internacional
de Medicina Veterinaria Preventiva y Enfermedades Exticas, La Habana, Cuba,
1987.
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
32). DE ARMAS, R.; R. SOLANO y J.R CARAL: Transferencia no quirrgica de
embriones en el ganado bovino. Resmenes Vil Jornada Cientfica para el
Mejoramiento Animal (CIMA), p. 55,1985.
33). IGLESIAS, C.; R.SOLANO y J.R CARAL: Trasplante de embriones en el ganado
bovino, fley. Cub. Reprod. Anim., 5: 33, 1979.
34). NEWCOMB, R.: Influence of synchrony and method of synchronization of results.
Monograph No. 16, Departament of Agriculture pp. 28-29, Canad, 1977.
35). PALMA, GA.: Recoleccin de embriones. En: Transferencia de embriones y
biotecnologa de la reproduccin en la especie bovina. Editorial Hemisferio Sur,
Argentina, 1993.
36). SOLANO, R.: Algunos aspectos para la evaluacin de embriones previo al
trasplante. En; Transferencia de embriones. CIMA, La Habana, 1988.
37). MARTAL, J.: Les biotechnologies de la reproduction animales. En: Biotechnologie,
4th. ed, (Ed.) Ren Scriban, Editorial Technique y Documentation, Lavoisier, Paris,
1993.
38). DE ARMAS, R.: Congelacin de embriones. Boletn de Reseas, Mejoramiento
Animal, Centro de Investigacin para el Mejoramiento Animal, La Habana, 1985.
39). PALMA, G.A. y G. BREM: Transferencia de embriones y biotecnologa de la
reproduccin en la especie bovina. Editorial Hemisferio Sur, Argentina, 1993.
40). CROZET, N.: Fertilization in vivo and in vitro. En: production in mammals and
man. (Eds.) C. Thibault; M.C. Levasseur and R.H.F. Hunter, Ellipses, Paris, 1993.
41). PREZ PREZ, F: El fascinante avance de las biotecnologas. Discurso para la
recepcin pblica como acadmico electo. Instituto de Espaa, Real Academia
Nacional de Medicina, Madrid, 1990.
42). BERG, U. y H.D. REICHNBACH: Produccin in Vitro de embriones. En:
Transferencia de embriones y biotecnologa de la reproduccin en la especie bovina.
Editorial Hemisferio Sur, Argentina, 1993.
43). NICOLS, J.F. and C. DELOUIS: Transgenic animals. Genetic manipulation of the
germ line. En: Reproduction in mammals and man. (Eds.) C. Thibault; M.C.
Levasseur and R.H.F. Hunter, Ellipses, Paris, 1993.
44). BREM, G.; U. BERG y H.D. REICHENBACH: Produccin de animales
transgnicos. En: transferencia de embriones y biotecnologa de la reproduccin en
la especie bovina. Editorial Hemisferio Sur, Argentina, 1993.
Catedrtica: compilador Dr. ngela Caldern
Universidad Estatal de Bolvar (FAC. CC. AGROP) MVZ
Catedrtica: compilador Dr. ngela Caldern
You might also like
- Operaciones auxiliares en reproducción ganadera. AGAX0108From EverandOperaciones auxiliares en reproducción ganadera. AGAX0108No ratings yet
- Comportamiento productivo y reproductivo en caprinos en trópico secoFrom EverandComportamiento productivo y reproductivo en caprinos en trópico secoRating: 1 out of 5 stars1/5 (1)
- Anatomia y Fisiologia Reproductiva de La Hembra BovinaDocument11 pagesAnatomia y Fisiologia Reproductiva de La Hembra Bovinaers55No ratings yet
- BovinosDocument93 pagesBovinosLeo Scott Pecho RojasNo ratings yet
- Consumo de Materia Seca en Bovinos LecherosDocument14 pagesConsumo de Materia Seca en Bovinos LecherosRafael CentenoNo ratings yet
- Manual de Manejo de Caprinos - Corredor Seco NicaraguaDocument21 pagesManual de Manejo de Caprinos - Corredor Seco NicaraguaAcf-e Misión Centroamérica0% (1)
- Rastro BovinosDocument13 pagesRastro BovinosDark ElfsNo ratings yet
- Inta Tecnica de Necropsia en Rumiantes 2019 Canton Odriozola PDFDocument49 pagesInta Tecnica de Necropsia en Rumiantes 2019 Canton Odriozola PDFFlorencia Mattío100% (1)
- Consanguinidad 2020Document80 pagesConsanguinidad 2020Lebni IdrovoNo ratings yet
- Parasitologia VeterinariaDocument7 pagesParasitologia VeterinariaYonairo HerreraNo ratings yet
- Inseminacion Artificial ZGralDocument32 pagesInseminacion Artificial ZGralCritica Perú Nadie SE SalvaNo ratings yet
- Leptospira en BovinosDocument7 pagesLeptospira en BovinosShellGarciaNo ratings yet
- Clase III Gestacion Implantacio N y Placentacion. Diag Gest. 2.0Document61 pagesClase III Gestacion Implantacio N y Placentacion. Diag Gest. 2.0tomas contrerasNo ratings yet
- Facultad Zootecnia PDFDocument69 pagesFacultad Zootecnia PDFLuis-mi Champi Gutierrez50% (2)
- Razas CebuinasDocument24 pagesRazas CebuinasFelipe MendozaNo ratings yet
- Enfermedades Reproductivas de Los BovinosDocument9 pagesEnfermedades Reproductivas de Los BovinosLight DarkNo ratings yet
- Palpacion RectalDocument130 pagesPalpacion RectalAnonymous QNfesIQxNo ratings yet
- Diagnostico de Gestacion Por Palpacion Rectal en BovinosDocument12 pagesDiagnostico de Gestacion Por Palpacion Rectal en Bovinosprome7eo100% (2)
- Morfometria en OvinosDocument8 pagesMorfometria en OvinosKarla Jacqueline Viveros RiofrioNo ratings yet
- Clase 1 de ZootecniaDocument12 pagesClase 1 de ZootecniaJohan UsecheNo ratings yet
- Queratoconjuntivitis Infecciosa BovinaDocument6 pagesQueratoconjuntivitis Infecciosa BovinaJaime Francisco Gómez AbregoNo ratings yet
- Raza HerefordDocument13 pagesRaza HerefordedijamesNo ratings yet
- Sistemas de Produccion AnimalDocument24 pagesSistemas de Produccion AnimalAndrea Santamaria100% (1)
- Cunicultura PDFDocument29 pagesCunicultura PDFJose BarriosNo ratings yet
- La Funcion Reproductiva Del Ganado BovinoDocument4 pagesLa Funcion Reproductiva Del Ganado BovinoPortal Ganadero100% (3)
- Texto Epidemiología y EpizootiologíaDocument17 pagesTexto Epidemiología y Epizootiologíafernie3007No ratings yet
- Evaluación de La VacaDocument118 pagesEvaluación de La VacaJUANSANESNo ratings yet
- Monografia de Anaplasmosis y PiroplasmosisDocument31 pagesMonografia de Anaplasmosis y PiroplasmosisKatty Zanabria100% (1)
- Anatomia y Fisiologia de Los RumiantesDocument37 pagesAnatomia y Fisiologia de Los Rumiantesdaddy rey0% (1)
- Vademecum ADLER Farma Full LNKDocument80 pagesVademecum ADLER Farma Full LNKAmadeo Bustos PonceNo ratings yet
- Manual de ZootecniaDocument4 pagesManual de ZootecniaLeonardo FernandezNo ratings yet
- Reducción de Hernia Umbilical en Bovinos: MVZ, Epa, Erb Antonio Flores PerezDocument24 pagesReducción de Hernia Umbilical en Bovinos: MVZ, Epa, Erb Antonio Flores PerezGregorio Emmanuel Contreras AltamiranoNo ratings yet
- Inseminacion Artificial de EquinosDocument47 pagesInseminacion Artificial de EquinosMafer CedeñoNo ratings yet
- Enfermedades Parasitarias de Las AvesDocument86 pagesEnfermedades Parasitarias de Las AvesMishell PrietoNo ratings yet
- Ciclo Estral en La OvejaDocument30 pagesCiclo Estral en La OvejaluzmarinaNo ratings yet
- Digestivo MonogastricosDocument43 pagesDigestivo MonogastricosMvz Mary Cielo Fernández Fuentes100% (1)
- Clase de PorcinosDocument81 pagesClase de PorcinosLesly Huarcaya CondoriNo ratings yet
- Enfermedades en Los Conejos PDFDocument40 pagesEnfermedades en Los Conejos PDFHans Velasco100% (2)
- Calendario Manejo AlpacasDocument24 pagesCalendario Manejo AlpacasmaricruzNo ratings yet
- Examen Clínico en Bovinos LecherosDocument14 pagesExamen Clínico en Bovinos LecherosVinicio Rodriguez MirandaNo ratings yet
- Toxocara CanisDocument54 pagesToxocara CanisPadrinoxD100% (2)
- Caprinos y OvinosDocument15 pagesCaprinos y Ovinoskarly echeverryNo ratings yet
- Alimentación CabrasDocument10 pagesAlimentación CabrasIsamar RojasNo ratings yet
- Inseminacion Artificial en OvinosDocument25 pagesInseminacion Artificial en OvinosDiego García MedinaNo ratings yet
- Enfermedades Ovinos Sistemas Cria IntensivaDocument17 pagesEnfermedades Ovinos Sistemas Cria IntensivaMauricio PeñaNo ratings yet
- Inseminación Artificial OvinosDocument5 pagesInseminación Artificial OvinosJuan José Aparicio PorresNo ratings yet
- Razas de BovinosDocument75 pagesRazas de BovinosJefrey HernandezNo ratings yet
- Medicina Preventiva BovinosDocument78 pagesMedicina Preventiva BovinosKarlichis Martinez50% (4)
- Cronometria DentariaDocument32 pagesCronometria DentariaNorlan Ariel Caldera NavarreteNo ratings yet
- Listeriosis BovinaDocument15 pagesListeriosis BovinaAlex Amaya50% (2)
- Manual Sanidad AnimalDocument275 pagesManual Sanidad AnimalKevyn AlejosNo ratings yet
- Razas Lecheras y Doble Propósito EuropeasDocument30 pagesRazas Lecheras y Doble Propósito Europeasestefany vertelNo ratings yet
- Aptitud Reproductiva Potencial y FuncionalDocument32 pagesAptitud Reproductiva Potencial y Funcionalfrank1049100% (1)
- Principales Enfermedades de RumiantesDocument88 pagesPrincipales Enfermedades de RumiantesCristhian Eduardo Poletti SchremppNo ratings yet
- Guia Final de BovinosDocument497 pagesGuia Final de Bovinosashlly mora100% (2)
- Anatomía y Fisiología de La Reproducción BovinaDocument5 pagesAnatomía y Fisiología de La Reproducción Bovinaafrich81No ratings yet
- Sistemas de Explotación BovinaDocument18 pagesSistemas de Explotación BovinaBenjamin Santos SalinasNo ratings yet
- Los animales domésticos criollos y colombianos en la producción pecuaria nacionalFrom EverandLos animales domésticos criollos y colombianos en la producción pecuaria nacionalNo ratings yet
- UF2168 - Control de incorporación y sanitario de los animales de recría y ceboFrom EverandUF2168 - Control de incorporación y sanitario de los animales de recría y ceboRating: 2.5 out of 5 stars2.5/5 (3)
- Leguminosas espontaneas de posible valor forrajero en ColombiaFrom EverandLeguminosas espontaneas de posible valor forrajero en ColombiaNo ratings yet
- Manejo de CampoDocument26 pagesManejo de CampoAlecsander RomeroNo ratings yet
- Materia Patologia ACTUALDocument51 pagesMateria Patologia ACTUALAlecsander RomeroNo ratings yet
- Ficha Estetica CorporalDocument7 pagesFicha Estetica CorporalAlecsander RomeroNo ratings yet
- Niveles Inec 2018Document6 pagesNiveles Inec 2018Alecsander RomeroNo ratings yet
- Licencia AntivirusDocument2 pagesLicencia AntivirusAlecsander RomeroNo ratings yet
- Basf Opera 2015Document2 pagesBasf Opera 2015Alecsander RomeroNo ratings yet
- Formato CuyesDocument2 pagesFormato CuyesAlecsander RomeroNo ratings yet
- Sistema Sanguineo VeterinariaDocument33 pagesSistema Sanguineo VeterinariaAlecsander RomeroNo ratings yet
- El EquidnaDocument7 pagesEl EquidnaAlecsander RomeroNo ratings yet
- Anatomía y Fisiología Del CorazónDocument14 pagesAnatomía y Fisiología Del CorazónAlecsander RomeroNo ratings yet
- Ejercicio de PorcinosDocument12 pagesEjercicio de PorcinosAlecsander RomeroNo ratings yet
- Sistema Reproductor FemeninoDocument9 pagesSistema Reproductor FemeninoSalmeron Herbert0% (1)
- Equipo 5. Hernia InguinalDocument8 pagesEquipo 5. Hernia InguinalLizbeth NotarioNo ratings yet
- 01a Clase Ginecología Anatomia Genital FemeninaDocument59 pages01a Clase Ginecología Anatomia Genital FemeninaEvanns Vela RuizNo ratings yet
- Maternidad SubrogadaDocument12 pagesMaternidad SubrogadaEduardo MalfavonNo ratings yet
- Proceso Embarazo - Parto - PuerperioDocument31 pagesProceso Embarazo - Parto - PuerperioJazmin ArandaNo ratings yet
- El PartoDocument187 pagesEl PartoAlexandra DelgadoNo ratings yet
- CARATULADocument4 pagesCARATULAYerko Flores MamaniNo ratings yet
- ADENOMIOSISDocument14 pagesADENOMIOSISDra Miozoti MendezNo ratings yet
- CERDOSDocument171 pagesCERDOSAlejandra María Castañeda ArcilaNo ratings yet
- Trabajo de Sexualidad HumanaDocument55 pagesTrabajo de Sexualidad HumanameissyNo ratings yet
- Modificaciones Fisiológicas de La Mujer Durante El EmbarazoDocument14 pagesModificaciones Fisiológicas de La Mujer Durante El EmbarazoFelipe GallardoNo ratings yet
- Reproductor Femenino (Histología y Anatomía)Document9 pagesReproductor Femenino (Histología y Anatomía)JatnielNo ratings yet
- Prctica 1 Aparatos Reproductores en VertebradosDocument5 pagesPrctica 1 Aparatos Reproductores en Vertebradosapi-261817550No ratings yet
- Materno Infantil 1 GlosarioDocument44 pagesMaterno Infantil 1 GlosarioSarmiento Derwin50% (2)
- Anatomia Del Sistema Reproductor FemeninoDocument31 pagesAnatomia Del Sistema Reproductor FemeninoLeidy Villamizar100% (2)
- Ejemplo de Record OperatorioDocument4 pagesEjemplo de Record OperatorioDean Ortiz Lopez50% (2)
- Viabilidad de Los GametosDocument5 pagesViabilidad de Los GametosCarol Ríos NúñezNo ratings yet
- Amenaza de AbortoDocument3 pagesAmenaza de AbortoAna maria maduroNo ratings yet
- Anatomia Gincologica..Dra CentellasDocument5 pagesAnatomia Gincologica..Dra CentellasRolando QuirozNo ratings yet
- Ciclos Sexuales PDFDocument20 pagesCiclos Sexuales PDFJesús AnguloNo ratings yet
- Esquema Tesis - Uap GudyDocument71 pagesEsquema Tesis - Uap GudyFredy Isidro Pariona RiveraNo ratings yet
- HISTERECTOMIA (Autoguardado)Document8 pagesHISTERECTOMIA (Autoguardado)Evelyn Karolina CuviNo ratings yet
- Cancer Apuesta Por La Vida LibroDocument126 pagesCancer Apuesta Por La Vida Librooezaki1No ratings yet
- ArfDocument29 pagesArfAlejandra MojicaNo ratings yet
- Historia Clínica GinecológicaDocument39 pagesHistoria Clínica GinecológicaCinthia CordovaNo ratings yet
- Cómo Comienza Un EmbarazoDocument5 pagesCómo Comienza Un EmbarazoVanessa Ramirez OjedaNo ratings yet
- Tercera RotacionDocument20 pagesTercera RotacionMariela Mina RiveraNo ratings yet
- Rotafolio Prev CACU y PAPDocument26 pagesRotafolio Prev CACU y PAPSantiago La TorreNo ratings yet
- 1°-2° GinecologiaDocument342 pages1°-2° GinecologiaZack Montoya BetancourtNo ratings yet
- Patologia de Utero y EndometrioDocument16 pagesPatologia de Utero y EndometrioJorge ReyesNo ratings yet