Professional Documents
Culture Documents
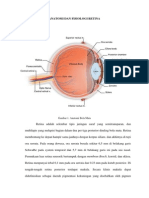
Anatomi Retina Dan Viterus
Uploaded by
riskiamanda13Copyright
Available Formats
Share this document
Did you find this document useful?
Is this content inappropriate?
Report this DocumentCopyright:
Available Formats
Anatomi Retina Dan Viterus
Uploaded by
riskiamanda13Copyright:
Available Formats
BAB I
PENDAHULUAN
Retina merupakan membran yang sangat tipis dan transparan. Ketertarikan
untuk mengenal anatomi retina akhir-akhir ini semakin besar karena pentingnya
fungsi retina terhadap sistem penglihatan. Retina pertama kali diperkenalkan oleh
Herophilus of Chalcedon (300 tahun SM), nama retina diberikan oleh Rufos of
Ephesus (110 M). Dinamakan demikian oleh karena penampakannya yang
berbentuk jala yang menangkap vitreus. Meskipun pada awalnya Galens
menggambarkan adanya kemiripan retina dengan otak, dimana perhatian lebih
menekankan pada vaskularisasi dan hubungannya dengan nervus optik, namun
Kepler (1608) pada akhirnya memperkenalkan retina sebagai jaringan fotoreseptor
primer pada mata. Pada akhir tahun 1600, Antoni van leeuwanhoek
menggambarkan sel globular dan pembuluh darah pada retina bovine.1
Korpus vitreous menempati sekitar 80% dari volume bola mata yaitu sekitar
4 ml dengan berat 4 g dan berat jenis 1,0053-1,0089. Pada bagian anterior korpus
vitreus berbatasan dengan korpus ciliaris, zonula,dan lensa,sedangkan pada bagian
posterior berbatasan dengan retina. Korpus vitreus memiliki beberapa fungsi
fisiologis, namun sebagai dasar untuk memahami fisiologi dan patofisiologi dari
korpus vitreus tersebut kita harus memahami anatomi, biokimiawi dan biofisika
dari korpus vitreus.2,3,4,5
Pemeriksaan dari korpus vitreus baik struktur maupun fungsinya dihambat
oleh dua hal yang sangat mendasar sehingga menyulitkan untuk memahami
fisiologi dan patofisiologi dari korpus vitreus tersebut. Yang pertama, adanya
kesukaran dalam memvisualisasikan morfologi dari korpus vitreus mengingat
1
strukturnya yang invisible. Yang kedua, berbagai tehnik yang telah dicoba untuk
menggambarkan struktur dari korpus vitreus yang dikombinasi dengan berbagai
artifact
ternyata
memberikan
interpretasi
yang
sulit
untuk
dibuktikan
kebenarannya secara invivo.2
Dalam sari pustaka ini akan dibahas lebih lanjut mengenai embriologi,
anatomi, dan fisiologi dari Retina dan korpus vitreus.
BAB II
TINJAUAN PUSTAKA
A. ANATOMI RETINA
Retina merupakan jaringan transparan yang melekat pada dinding
posterior bola mata. Retina melebar dari makula di posterior hingga pada sekitar 5
mm dari ekuator anterior yakni ora serrata dimana jaringan retina menyatu
dengan epitel tak berpigmen dari pars plana silia. Jaringan retina melekat longgar
dengan lapisan RPE dibawahnya dan dapat dengan mudah dipisahkan pada
specimen postmortem. Retina melekat kuat pada daerah diskus optikus dan ora
serrata. Retina juga melekat pada vitreus base.1
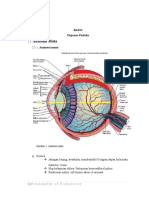
Topografi Retina
Ketebalan retina bervariasi pada setiap bagian, sekitar 0,1 mm 0,5 mm.
Hal ini sangat penting diketahui dalam aplikasi klinis.6
1. Area sentralis-Makula
Macula lutea atau bintik kuning merupakan bagian dari retina yang banyak
mengandung pigmen xantophil atau pigmen kuning. Daerah macula, secara
histologis digambarkan sebagai area yang terdiri atas 2 atau lebih lapisan ganglion
dengan diameter 5-6 mm dan berada ditengah antara arcade vascular nasal dan
temporal. Makula lutea 1 mm ke lateral, 0.8 mm ke atas dan di bawah fovea, 0.3
mm dibawah meridian horizontal serta 3.5 mm ke arah tepi nervus optik.6
2. Fovea
Daerah sentral dari macula, berukuran 1,5 mm di sebut sebagai fovea atau
fovea sentralis, yang secara anatomis dan komposisi sel fotoreseptornya
merupakan daerah untuk ketajaman penglihatan dan penglihatan warna. Daerah
ini memiliki tingkat kepadatan sel cones tertinggi, yakni mencapai 143.000/mm 3.
Didalam fovea terdapat daerah yang tidak memiliki vaskularisasi, jadi dipelihara
oleh sirkulasi koriokapiler, yang disebut fovea avascular zone (FAZ). Secara klinis
dapat terlihat pada angiografi fluorosensi. Pada bagian tengah fovea di kenal
sebagai foveola, berukuran diameter 0.35 mm daerah yang berisi sel sel cone
ramping yang tersusun rapat.6
3. Parafovea
Di sekitar lingkaran fovea, terdapat area dengan lebar sekitar 0.5 mm dan
diameter total sekitar 2.5 mm disebut area parafoveal. Mengandung akumulasi
neuron terbesar, terdapat lapisan sel ganglion, lapisan inti dalam, dan lapisan
pleksiform luar yang tebal. Di daerah ini pula lapisan plexiform luar mengalami
penebalan, yang disebut lapisan Henle, dibentuk oleh berlapis-lapis axon
fotoreseptor dari foveola. Pada bagian ini sudah mulai terlihat adanya rods 1,6
4. Perifovea
Diluar zona tersebut terdapat lingkaran dengan ukuran 1.5 mm yang kenal
dengan perifoveal zone, merupakan lingkaran terluar dari area sentralis. Daerah
ini dimulai pada titik dimana lapisan sel ganglion mulai memiliki empat baris
nucleus dan berakhir diperifer dimana sel ganglion hanya terdiri dari satu lapis
sel. Dari pemeriksaan funduskopi, daerah perivofea merupakan lingkaran dengan
lebar 1,25- 2,75 mm dari foveola, dengan diameter horizontal 5.5 mm. Daerah
perifovea ini berbeda dengan parafovea dikarenakan daerah ini memiliki sel
kepadatan sel cones yang jarang. 1
5.
Diskus
optik
Nervus
optik
meninggalkan retina sekitar 3 mm di sebelah medial makula lutea, tepatnya pada
diskus optik. Bagian tengah dari diskus optik sedikit terdepresi, dimana daerah ini
ditembus oleh arteri dan vena retina sentralis. Pada diskus optik sama sekali tidak
terdapat sel rod maupun sel cone, oleh karena itu
terhadap rangsangan
cahaya
daerah ini tidak sensitif
dan disebut blind spot. Pada pemeriksaan
funduskopi, diskus optik terlihat sebagai daerah berwarna pink pucat, lebih pucat
dari daerah di sekitarnya.7
6. Ora Serrata
Merupakan daerah perbatasan retina. Ditandai dengan persambungan antara
beberapa lapis pars optic retina dengan satu lapis epitel non pigmen korpus
siliaris. Karakteristik yang menonjol dari area ini adalah lapisannya yang tipis,
kurang vaskularisasi dan hubungan yang rapat dengan vitreus base dan zonula
fibers. Dinamakan ora serrata karena banyaknya takikan yang dibentuk oleh
elongasi jaringan retina kearah epitel siliaris.(1)
Retina terdiri atas 2 lapisan utama, yaitu lapisan Retina Pigment Epithelium
(RPE) di bagian luar, dan lapisan neurosensori dibagian dalam.1,7
A. Retinal Pigment Epithelium (RPE)
RPE adalah selapis sel- sel hexagonal yang tersebar dari diskus saraf optik
sampai ke ora serrata dimana lapisan ini berbatasan dengan epitel non pigmen
dari badan siliar. Strukturnya disesuaikan
dengan fungsinya, yaitu dalam
metabolisme vitamin A, menyeimbangkan sawar darah retina bagian luar,
fagositosis
segmen luar fotoreseptor,
basalis, produksi
pertukaran panas, membentuk lamina
matriks polisakarida yang mengelilingi segmen luar dan
berperan dalam transport aktif materi- materi yang masuk dan keluar dari RPE.7
Seperti sel epitel dan endotel lainnya, sel- sel RPE juga terpolarisasi.
Permukaan basalnya berlekuk- lekuk dan menyediakan permukaan yang luas
sebagai tempat melekatnya lamina basalis yang membentuk lapisan dalam dari
membran Bruch. Apeksnya mempunyai tinjolan vili- vili yang berbatasan dengan
segmen luar lapisan fotoreseptor, ditautkan oleh matriks mukopolisakarida
(matriks inferoreseptor) yang mengandung kondroitin -6- sulfat, asam sialat dan
asam hyaluronat. Terpisahnya lapisan RPE dan lapisan neurosensori retina disebut
ablasi retina.7
Gambar 7.
Retinal Pigmen
Epithelium
(RPE)
(13)
Sel sel RPE melekat satu dengan lainnya melalui pertautan interseluler
kompleks. Zonula okludens dan zonula adheren tidak hanya berfungsi
mempertahankan bentuk dan stabilitas dari struktur RPE, tetapi juga memainkan
peranan penting dalam menjaga keseimbangan sawar darah retina bagian luar.
Zonula okludens terdiri dari membran plasma yang bersatu membentuk pita
sirkular atau sabuk antara satu sel dengan yang lain. Pada ruang interseluler
terdapat zonula adherens.7
Sel- sel neurosensori dan sel RPE memiliki perbedaan penting pada daerahdaerah tertentu. Sel- sel neurosensori paling tebal pada daerah papillomacular
bundle dekat dengan saraf optik (0.23 mm) dan paling tipis pada foveola (0.10
mm) dan ora serrata (0.11 mm). Sel- sel RPE mempunyai diameter yang
bervariasi antara 10-60 um. Dibandingkan dengan sel- sel RPE yang terletak di
daerah perifer, sel- sel RPE di fovea lebih tinggi dan lebih tipis serta mengandung
melanosom yang lebih banyak dan lebih besar. Sel sel RPE yang terletak
diperifer lebih pendek, lebar dan kurang mengandung pigmen. Tidak terlihat
proses mitosis dari sel- sel RPE pada mata dewasa normal.7
Sitoplasma dari sel- sel RPE mengandung granula-granula pigmen yang
bulat dan oval (melanosom). Organel- organel ini berkembang selama
pembentukan optic cup dan terlihat pertama kali sebagai non melanin
premelanosom. Sitoplasma dari sel-sel RPE juga mengandung mitokondria ,
reticulum endoplasma, apparatus golgi dan sebuah nucleus yang
bulat dan besar.
Seiring dengan pertambahan usia, badan sisa yang yang tidak terfagositosis
sempurna, pigmen lipofuchsin, fagosom dan material- material lain akan
diekskresikan oleh RPE dibawah lamina basalis RPE. Hal ini yang menyebabkan
terbentuknya drusen. Drusen berada diantara membrana basalis RPE dengan zona
kolagen membrana Bruch.
B. Neurosensori Retina
pembentuknya
Gambar 8. Lapisan retina dan komponen
Membrana limitans eksterna
Lapisan ini merupakan membran tipis, kumpulan intermediate junction
zonula adherens antara segmen dalam rod dan cone dengan apex sel- sel Muller ,
serta sel Muller satu dengan lainnya. Pertautan ini merupakan batas ruang
subretinal dam menjadi barrier dari difusi molekul besar kedalan maupun keluar
ruangan subretinal.8
Lapisan nuklear luar
Merupakan lapisan yang ditempati oleh nukleus dan badan sel rod dan
cones. Di daerah parafovea lapisan ini mengandung 8-10 lapisan
nucleus
terutama dimiliki oleh cone.8
Lapisan pleksiform luar
Lapisan ini terdiri dari sinaps rod dan cone dengan dendrite sel horizontal
dan sel bipolar. Sinaps dari Rod berbentuk oval dan disebut spherules, sedangkan
terminal cones lebih lebar disebut pedicles. Keduanya berinvaginasi ke dalam
terminal fotoreseptor dan keduanya dipisahkan oleh jembatan presinaps dan
sinaptic ribbon. Lapisan pleksiform luar paling tebal didaerah makula, berisi
akson dari rod dan cones tersusun radial dari fovea, yang merupakan lapisan serat
Henle.6
Lapisan nuklear dalam
Lapisan nuklear dalam terdiri atas 4 jenis sel, yaitu : sel bipolar, sel
horizontal, sel amakrin. Sel horizontal berada pada bagian distal dari lapisan
nuklear dalam, sementara sel amakrin terletak di bagian paling proksimal.
Nukleus sel bipolar terletak pada bagian intermediate luar dan intermediate dalam
dari lapisan ini.8
Lapisan pleksiform dalam
Sinaps pada lapisan pleksiform dalam jauh lebih kompleks dibanding
dengan lapisan pleksiform luar. Ketebalannya bervariasi antara 18 dan 36 m dan
tidak terdapat pada daerah foveola. Terminal sel bipolar dan dendrit dari sel
amakrin dan sel ganglion saling berhubungan pada level yang berbeda di lapisan
pleksiform dalam.8
Lapisan sel ganglion
Lapisan ini berisi badan sel dari sel-sel ganglion, jumlah sel ganglion
ditemukan sekitar 0,7- 1,5 juta pada retina orang muda, di lapisan ini juga terdapat
sel amakrin, didaerah perifer retina hanya terdapat 1 baris sel-sel ganglion tetapi
didalam macula terdapat 10 baris sel-sel ganglion, dengan konsentrasi tertinggi
10
didaerah parafovea. Masing-masing sel ganglion menerima sinaps melalui
dendritnya didaerah pleksiform dalam dan mengirimkan akson ke lapisan serat
saraf.8
Lapisan serabut saraf
Lapisan ini terdiri dari akson sel- sel ganglion, yang berasal dari seluruh
bagian retina kemudian menuju ke diskus optik untuk membentuk nervus optic.
Kumpulan akson ini akan dikelilingi oleh Muller dan astroglial. Lapisan ini paling
tebal di daerah diskus optik, yaitu sekitar 20-30 m dan paling tipis di perifer.
Membrana limitans interna
Lapisan ini bukan merupakan membran sepenuhnya. Lapisan ini dibentuk
oleh end footplate sel Muller dan perlekatan dengan lamina basalis. Tebalnya
sekitar 1-2 m. Membran ini bersatu dengan fibril kolagen vitreus.8
Elemen - Elemen Neurosensori Retina
Rods
Used for night vision
Very light sensitive; sensitive to
Cones
Used for day vision
At least 1/10th of the rods light
scattered
sensitive;sensitive only to direct
light
(have
more
pigment than cones)
Loss causes night blindness
Low visual acuity
light
Loss causes legal blindness
High visual acuity; better spacial
Not present in fovea
Slow response to light, stimuli
resolution
Concentrated in fovea
Fast
response to light, can
added over time
perceive more rapid change in
Stacks of membrane-enclosed
stimuli
Disks are
disks are unattached to cell
membrane
attached
to
outer
of
photosensitive
pigment (monochrome vision)
Confer achromatic vision
1. Elemen neuronal
Perbandingan
struktur sel
rod dan cones
Tabel
4. Perbedaan
struktur Rod
membrane
20 times more rods than cones in
the retina
One type
Gambar 5.
dan cones
Three
types
of
photosensitive
pigment in human (color vision)
Confer color vision
11
Lapisan fotoreseptor mengandung neuroepithel khusus yaitu sel rods dan
cones. Setiap sel fotoreseptor ini mempunyai segmen luar dan segmen dalam.
Segmen luar dikelilingi oleh matriks mukopolisakarida yang menyebabkan kontak
antara sel fotoreseptor dan tonjolan vili dari RPE. Tight junction atau penghubung
interseluler lainnya tidak terdapat diantara segmen luar sel fotoreseptor dengan
RPE, sehingga memungkinkan untuk terjadinya transport aktif dari RPE ke sel
fotoreseptor, namun juga berarti tidak adanya penunjang yang menjaga posisi dari
pertautan sel- sel ini.8
Sel rod mempunyai segmen luar yang mengandung keping- keping disk
menyerupai koin dan sebuah silium perantara. Segmen dalam dari sel rod terbagi
menjadi dua elemen yaitu berada diluar berbentuk elips mengandung banyak
mitokondria dan yang berada didalam berbentuk myoid mengandung banyak
glikogen. Elemen myoid ini berlanjut dengan badan sel utama dimana terdapat
nucleus. Bagian paling dalam dari sel rod ini mempunyai badan sinaptik atau
spherule, dimana struktur ini terbentuk dari invaginasi tunggal yang
mempetemukan dua tonjolan sel horizontal dan satu atau lebih dendrit sel bipolar.8
Sel cones ekstrafovea juga mempunyai elemen berbentuk elips dan myoid.
Nucleus dari sel cone ini lebih dekat dengan membrana limitans eksterna
dibanding sel rod.1
Meskipun secara garis besar struktur segmen luar dari sel rod dan cone
memiliki kesamaan, namun setidaknya ada satu perbedaan penting. Lempeng
diskus dari rod tidak melekat pada membran sel, lempeng diskus ini merupakan
suatu struktur tersendiri. Lempeng diskus dari cones melekat pada membran sel.
Badan sinaptik dari cone atau pedikel lebih kompleks dibanding spherule dari rod.
Pedikel cone bersinaps dengan sel- sel cones lainnya, dengan sel- sel rod beserta
sel bipolar dan horizontal.1
12
Gambar 10. Segmen luar sel rod
dan sel cone
Rod spherule dan Cone pedicle
Prosesus dari sel bipolar dan sel horizontal berinvaginasi ke rod spherule,
dimana ketiga komponen ini membentuk struktur yang disebut Triad, yang
biasanya satu triad untuk satu spherule. Rod spherule terdiri dari mitokondria,
mikrotubulus dan neuron presinaptik. Celah sinaptik yang kecil memisahkan sel
presinaptik dan membran postsinaptik. Pada bagian dalam spherule terdapat
ribbon. Antara ribbon dan membran sel terdapat arcuate density. Ribbon dan
arcuate density dikelilingi oleh vesikel sinaptik dan berperan dalam pelepasan
neurotransmitter dari vesikel sinaptik. Conventional vascular neurotransmitter
membawa informasi dari sel fotoreseptor ke post sinaptik prosesus sel bipolar dan
sel horizontal.7
Struktur cone pedicle mirip dengan rod spherule, tetapi bentuknya lebih besar
dan terdiri dari beberapa triad, sampai sekitar 25. Ribbon sinaptiknya lebih kecil
dan lebih banyak. Midget bipolar cell mengirimkan seluruh dendritnya
bagian tengah cone triad.
ke
13
Gambar 11. Rod spherule dan cone pedicle di lapisan plexiform luar(11)
Gambar 12.A.
Gambar 12.B.
Pedikel
Spherule(11)
Rod
Cone
Dendrit dari sel bipolar lainnya dari basal atau superfisial mengadakan
kontak pada tiap bagian terminal invaginasi, yang kemudian disebut invaginating
midget bipolar cell dan flat midget bipolar cell.
Setiap cone pedicle memiliki
banyak invaginasi. Pada bagian sentral retina dimana pedikel lebih kecil sebanyak
15 sampai 25 invaginasi terdapat pada setiap terminal.7
INTERNEURONS
14
Sel bipolar, horizontal, dan sel amakrin disebut interneurons, dimana badanbadan sel ini membentuk lapisan inti dalam. Nukleus dari sel Muller juga
ditemukan dilapisan ini.6
Gambar 13. Skematik diagram tipe sel dan lapisan histologi retina(10)
Sel Bipolar
Sel bipolar membawa sinyal dari sel fotoreseptor ke sel ganglion atau sel
amakrin, terdapat 2 kelas utama sel bipolar, yaitu Rod bipolar cells, yang
berhubungan dengan spherule rod dan Cone bipolar cells dan yang berhubungan
dengan pedikel cone, terdiri atas midget cone bipolar cells dan diffuse cone
bipolar cells.7
15
Gambar14. Sel bipolar (warna merah)(13)
Dendrit
dari
diffuse
cone
bipolar
cells
memberikan cabangnya ke lapisan pleksiform
luar
dan mengadakan kontak dengan beberapa
pedikel cone. Pada sisi yang berlawanan, akson
dari diffuse bipolar cells diproyeksikan ke lapisan pleksiform dalam dan
berhubungan dengan dendrite dari sel ganglion.
Midget cone bipolar cells
bersinaps dengan satu pedikel cone dan satu akson mengadakan kontak dengan
satu sel ganglion. Pada dasarnya, midget cone bipolar cells berhubungan dengan
satu sel cone ke serat saraf optik. Sebaliknya, diffuse bipolar cell mempunyai
lebih banyak jalur input dan output. Nucleus dari sel bipolar membentuk lapisan
inti dalam. Baik rod bipolar cells maupun cone bipolar cell menggunakan
glutamate untuk proses neurotransmisi.8
Sel Horizontal
Sel-sel horizontal (dan sel amakrin) dianggap sebagai local-circuit neurons.
Neurit sel horizontal berakhir pada pedikel cone. Satu buah cabang sinaps neurit
bersinaps baik dengan spherule rods maupun pedikel cone. Sinaps ini terjadi pada
lapisan plexiform luar dan distribusi aksonal mengindikasikan bahwa sel
horizontal berintegrasi dengan sel rod dan cone pada area yang berbatasan pada
retina.7
16
Gambar
15.
Sel
horizontal
Terdapat 2 tipe sel horizontal, yairu sel horizontal tipe 1 yang ditandai
dengan adanya dendrite yang besar yang mengadakan kontak hanya dengan sel
cones dan sebuah akson panjang yang berakhir pada terminal akson yang hanya
berhubungan dengan sel rod. Sel horizontal menggunakan GABA sebagai
neurotransmitter. Horizontal tipe 2 hanya kontak dengan cone dengan cabang
dendrite yang kecil serta axon yang pendek.6,7
Sel Amakrin
Gambar 16. Sel amakrin
Sebagian besar sel amakrin berlokasi pada bagian proksimal dari lapisan inti
dalam. Sel ini memodulasi sinyal pada lapisan pleksiform dalam. Sel amakrin
dapat diklasifikasikan berdasarkan dendritic field diameter menjadi : narrow
field(30-150um), small field (150-300 um) dan medium field (300-500 um).
17
Berdasarkan distribusi dendrit pada lapisan pleksiform dalam, sel amakrin dapat
diklasifikasikan sebagai stratified atau diffuse cell. Substansi neuroaktif yang
terdapat pada sel amakrin yaitu glisin, GABA, asetilkolin, serotonin, dopamine,
nitit oxide, neurotensin, enchepalin, somatostatin, substansi P, vasoactive
intestinal peptide(VIP), dan glucagon.
Sel Ganglion
Sel ganglion terletak diantara lapisan pleksiform dalam dengan lapisan
serabut saraf. Di daerah sekitar fovea sentralis terdapat 5-7 lapis sel- sel ganglion
dan merupakan lapisan sel ganglion yang paling tebal (80um) dan lapisan sel
ganglion paling tipis terdapat pada daerah perifer retina (10um). Terdapat satu sel
ganglion untuk setiap 100 sel rod dan satu sel untuk setiap 5 sel kerucut. Sel- sel
ganglion memiliki 2 jenis sinaps intersel. Dendritnya bersinaps dengan akson selsel bipolar dan amakrin.7
Akson-akson dari sel-sel ganglion melingkar untuk membentuk permukaan
dalam dari retina secara paralel, dimana akson- akson ini membentuk lapisan
serabut saraf yang nantinya bersatu pada diskus saraf optik.7
Setiap satu saraf optik memiliki lebih dari 1 juta serabut saraf. Serabutserabut saraf yang berasal dari temporal berjalan melengkung mengelilingi macula
untuk memasuki daerah superior dan inferior diskus saraf optik. Serabut- serabut
saraf papillomakular dan fovea berjalan lurus kedalam diskus saraf optik. Serabutserabut dari nasal berjalan secara radier. Serabut- serabut ini dapat dievaluasi
dengan menggunakan iluminasi sinar hijau (red free) pada oftalmoskop. 7
Elemen- elemen pada neuron pada
retina beserta koneksinya sangat
kompleks. Banyak tipe sel- sel bipolar, amakrin dan sel ganglion lain yang
berperan. Elemen- elemen neuron dimana lebih dari 120 juta sel rod dan 6 juta sel
cone saling berhubungan satu sama lain dan proses pengiriman sinyal antara
neurosensori retina sangat penting 7
2. Elemen Glial
Sel Muller
18
Gambar 17. skema sel Muller(10)
M : Sel muller, EF : End foot sel
muller, MV : Mikrovili sel muller
Sel- sel Muller adalah sel- sel glial yang berjalan secara vertikal dari
membran limitans eksterna menuju membran limitans interna. Nukleusnya berada
pada lapisan inti dalam. Sel- sel Muller, bersama elemen- elemen glial lainnya
(astrosit dan microglia) merupakan penunjang bagi retina.6
Beberapa studi terbaru memperlihatkan peranan akan pentingnya sel-sel
muller ini dalam perkembangan dan metabolisme retina. Imunohistokimia
memperlihatkan bahwa sel-sel ini mengandung retinaldehid binding protein ,
glutamine, taurin dan glutamine sythetase. Sel-sel muller juga berperan dalam
degradasi neurotransmitter GABA. Adanya RNA messenger yang mengkode
carbonic anhydrase II menunjukkan bahwa sel- sel ini juga penting dalam
menahan karbondioksida bebas yang dilepaskan ke ruang ekstraseluler oleh
elemen-elemen neurosensori retina. Produksi insulin dan faktor- faktor
pertumbuhan dari sel- sel muller ini juga sangat penting dalam metabolisme
retina.
Sel muller juga diduga menunjuang metabolisme neuron- neuron retina, oleh
karena sel muller ini merupakan tempat utama penyimpanan glikogen. Pada
kondisi stress metabolik yang tinggi, seperti hipoglikemia, pemecahan glikogen
19
pada sel muller ini dapat menyediakan metabolit- metabolit penting, seperti asam
laktat untuk digunakan oleh neuron yang rusak. Penelitian metabolisme glukosa
pada hewan coba menunjukkan bahwa hasil utama glikolisis adalah laktat.9
Fungsi lain dari Sel Muller adalah berperan dalam mempercepat adaptasi
gelap dari sel cones, peranannya diduga membantu RPE dalam siklus daur ulang
sel cones. Hal ini dibuktikan dengan percobaan pada hewan coba,
dengan
mengangkat lapisan RPE pada hewan tersebut, ternyata sel cones masih terus
dapat bekerja, peran RPE digantikan oleh sel- sel Muller ini.9
Astrocyte
Astrocyte tidak muncul dari neuroepithelium tetapi berimigrasi ke retina.
Permukaan astrocyte ditandai dengan adanya interaksi contac-spacing antar sel.6
Mikroglia
Mikroglia merupakan makrofag retina, berasal dari mesodermal, dari
monosit darah dan memasuki retina selama perkembangan pembuluh darah.
Ditemukan pada lapisan serabut saraf dan lapisan pleksiform dalam dan luar.6
B. FISIOLOGI RETINA
Kelangsungan fotoreseptor dan koriokapiler tergantung pada RPE. Jika
RPE mengalami gangguan, baik secara kimiawi maupun mekanik, maka
fotoreseptor dan koriokapiler akan mengalami atrofi. RPE memproduksi sitokin,
termasuk basic Fibroblast Growth Factor (bFGF) yang mempertahankan
kelangsungan fotoreseptor. Akan tetapi sampai saat ini masih belum jelas berbagai
senyawa yang dihasilkan oleh RPE dalam mendukung kelangsungan fotoreseptor
dan koriokapiler in vivo.
RPE berperan dalam memperbaharui segmen luar
fotoreseptor, penyimpanan dan metabolisme vitamin A, transport dan barrier
epitel. Selain itu juga mengabsorpsi cahaya oleh pigmen melanin di epitel,
menangkap redikal bebas oleh pigmen melanin, dan detoksifikasi obat oleh sistein
sitokrom P-450 retikulum endoplasmik halus yang juga ditemukan pada sel RPE.
(4)
20
FUNGSI IMUN
Fungsi lain dari retina adalah sebagai regulator respon imun lokal. Hal ini
dijalankan oleh barrier pasif RPE dan sekresi aktif sitokin imunosupresif seperti
TGF-. Saat terjadi inflamasi, maka RPE menghambat kerja mediator inflamasi.
RPE juga secara aktif mensekresi reseptor Tumor necrosis factor- (TNF-) . Sel
RPE memproduksi faktor yang mensupresi neutrophil superoxide generator yang
membatasi kerusakan jaringan selama terjadi inflamasi.6
RPE juga mengekspresikan Fas-ligand (FasL) yang digunakan untuk
proses apoptosis dalam menginvasi Fas-expressing yang diaktivasi oleh limfosit
atau sel endotelial neovaskular. RPE dianggap sebagai macrophage-like
morphology. Sel RPE secara aktif mengfagositosis dan menghambat reseptor
mediated phagositosis dari komponen matriks ekstraseluler.Proliferasi sel RPE
terjadi setelah stimulasi PDGF, TNF-, IGF dan VEGF. Aktivasi RPE bisa
mengekspresikan molekul yang berhubungan dengan pengikatan dan infiltrasi
lekosit. Terlihat pula ekspresi molekul-molekul adhesi seperti CD45, CD48,
CD54, CD59, CD68 dan ICAM-1. Saat diaktivasi oleh IL-1, TNF, IL-7 atau
glikasi albumin serum, maka sel RPE akan mengaktivasi lekosit dengan
mensekresi kemokin, yaitu monocyte chemotactic protein-1 (MCP-1) dan IL-8,
dimana kemokin ini bisa dihambat oleh dexamethasone dan cyclosporin.
BLOOD RETINAL BARRIER
Terdapat dua komponen pembentuk blood retinal barrier (BRB), yakni
inner blood retinal barrier dan outer blood retinal barrier. Integritas BRB telah
dikenali sebagai komponen penting dalam mempertahankan fungsi visual normal
dan gangguan pada BRB ini dapat memberikan manifestasi pada berbagai
penyakit, seperti macular edema, sebagai akibat kebocoran cairan albumin dan
protein, dengan akibat akumulasi eksudat intraretina.10
Outer blood retinal barrier dibentuk oleh RPE yang mengontrol pertukaran
cairan dan molekul antara koriokapiler yang berfenestra dan permukaan luar
retina. Dua komponen RPE yang berperan dalam fungsi barrier adalah tight
junction antara sel-sel RPE dan distribusi protein seluler yang terpolarisasi. Tight
21
junction merupakan struktur dinamik dimana permeabilitasnya dapat dipengaruhi
oleh kondisi fisiologis tertentu dan oleh agen-agen farmakologis. Regulasi
transport transepitelial bergantung pada distribusi protein seluler yang asimetris.6
FOTOTRANSDUKSI ROD
Proses penerimaan dan perubahan cahaya yang masuk kedalam retina
membutuhkan energi dimana respon retina ini membedakannya dengan struktur
saraf lainnya. Kombinasi dari proses ini melibatkan organel khusus dari sel
fotoresptor. Rod memiliki lebih banyak membran dibandingkan dengan cone
sehingga rod lebih sensitif.3
Segmen luar dari rod terutama mengandung material plasma membran yang
tersusun secara khas. Membran ini tersusun dalam bentuk kantung-kantung pipih
sepanjang aksis horizontal dari segmen luar. Terdapat sekitar 1000 kantung pada
segmen luar rod dan sekitar 1 juta molekul rhodopsin pada setiap kantung.
Kantung- kantung ini melayang- layang pada sitoplasma seperti sebuah tumpukan
koin yang tidak berhubungan dengan plasma membran luarnya. Kantung ini
mengandung mesin protein untuk menangkap dan memperkuat energi cahaya.
Melimpahnya membran sel luar ini meningkatkan jumlah molekul rhodopsin yang
dapat menyerap cahaya.3,6,8
Cahaya diserap oleh rhodopsin yang terletak pada membran sel luar dari rod.
Rhodopsin adalah sejenis protein berupa membran yang mudah ditembus dan
sejenis dengan reseptor alfa dan beta adrenergik. Setiap molekul bertanggung
jawab terhadap satu kuantum cahaya. Rhodopsin menyerap cahaya hijau dengan
panjang gelombang sekitar 510 nm. Rhodopsin kurang baik dalam menyerap
cahaya biru dan kuning dan tidak sensitif terhadap cahaya merah.6
Pada saat rhodopsin menyerap suatu kuantum cahaya, ikatan ganda dari IIcis retinal akan pecah dan molekul opsin mengalami perubahan konfigurasi yang
cepat, sehingga terjadi keadaan aktif yang disebut metarhodopsin II. Rhodopsin
yang terktivasi memulai reaksi dengan mengontrol aliran kation- kation kedalam
segmen luar rod. Target dari reaksi ini adalah pada pintu saluran cGMP (cyclic
Guaonosine Monophosphate) yang terletak di membran terluar dari segmen luar.
22
Saluran ini mengontrol aliran ion natrium dan kalsium kedalam rod. Dalam
suasana gelap, ion natrium dan kalsium mengalir melalui saluran ini dimana
terbukanya pintu saluran ini dipertahankanoleh cGMP. Keseimbangan ion
dipertahankan oleh pompa Na+, K+-ATPase pada segmen dalam dan Na+/K+- Ca
exchanger pada membran segmen luar, yang mana kedua proses ini membutuhkan
energi. Keadaan depolarisasi rod menyebabkan dilepasnya neurotransmitter
glutamate dari terminal sinaptik dan dimulailah sebuah sinyal neural dari proses
melihat.7
Gambar 20 . Aktivasi fototransduksi(11)
Rhodopsin yang telah teraktivasi merangsang molekul kedua, transdusin,
dengan cara menukar guanosin difosfat (GDP) dengan guanosin trifosfat (GTP).
Satu moloekul rhodopsin dapat mengaktifkan seratus molekul transdusin,
sehingga memperkuat reaksi. Transduksin yang aktif memicu protein ketiga, rod
fosfodiesterase (rod PDE) yang menghidrolisis cGMP ke 5-noncyclic GMP.
Penurunan cGMP ini menutup pintu saluran- saluran, dimana aakan menghentikan
masuknya natrium dan kalsium dan membuat keadaan hiperpolarisasi rod.
Hiperpolarisasi menghentikan pelepasan glutamate dari terminal sinaptik.7
Pada keadaan gelap, rod kembali ke keadaan gelapnya seiring dengan
terhentinya aliran reaksi. Rhodopsin mengalami inaktivasi akibat fosforilasi pada
23
C-terminalnya oleh rhodopsin kinase, yang dibantu oleh ikatan arrestin.
Transdusin dinonaktifkan akibat hidrolisis dari GTP ke GDP oleh aktivitas
GTPase transdusin intrinsic, yang juga menonaktifkan PDE. Guanilat siklase,
suatu enzim yang mensintesis cGMP dari GTP, diaktifkan oleh menurunnya kadar
kalsium intraseluler akibat tertutupnya saluran, aksi dari enzim ini dibantu oleh
protein pembantu-guanilat siklase (GCAPs). Dengan meningkatnya kadar cGMP,
pintu saluran- saluran tertutup dan rod kembali mengalami depolarisasi.
Meningkatnya kadar kalsium intraseluler mengembalikan aktivitas guanilat
siklase ke level gelapnya. Umpan balik kalsium juga dapat meregulasi fosforilasi
rhodopsin dengan jalan melindungi sensitivitas pintu saluran.7
FOTOTRANSDUKSI CONE
Fototransduksi yang terjadi pada cone adalah kebalikan dari rod. Cone-opsin
yang telah teraktivasi oleh cahaya memulai pengaliran enzimatik yang
menghidrolisis cGMP dan menutup saluran kation pada pintu spesifik cGMP cone
di membran segmen luarnya. Fototransduksi pada cone kurang sensitif namun
memiliki kemampuan yang cepat dalam beradaptasi terhadap berbagai kadar
iluminasi. Semakin besar kadar cahaya, maka semakin cepat dan akurat respon
dari cone. Kecepatan dan ketepatan sangat penting dalam kerja cone. Hal ini yang
menjadi alasan peningkatan ketajaman penglihatan seiring dengan peningkatan
iluminasi. Karena kemampuan cone dalam beradaptasi, cone sangat diperlukan
untuk ketajaman penglihatan.7
Pada cone, terdapat mekanisme umpan balik negatif. Sel-sel horizontal
bersinaps secara antagonis terhadap cone, dimana sel ini melepas GABA yang
bersifat inhibitor. Pada saat cahaya menghiperpolarisasi cone, maka cone
membuat hiperpolarisasi sel horizontal disebelahnya. Hal ini mengakibatkan
inhibisi terhadap sel horizontal, sehingga pelepasan GABA terhenti dan terjadi
depolarisasi cone. Keadaan depolarisasi ini menghambat keadaan hiperpolarisasi
oleh cahaya dan mencoba untuk mengembalikan cone pada keadaan
hiperpolarisasi oleh cahaya. Umpan balik negatif ini berfungsi agar cone tidak
mengalami keadaan overload (kelebihan beban) sehingga memungkinkan cone
dapat merespon stimulus baru dengan lebih cepat.6,7
24
PENGLIHATAN WARNA
Untuk melihat warna, mamalia harus memiliki sedikitnya 2 kelas spektral
berbeda dari cone. Pada mata manusia normal, ada 3 tipe sel cone dimana
ketiganya merupakan 3 sistem cone-opsin. 3 sistem cone-opsin tersebut adalah
short-wavelength sensitive (S), middle-wavelength-sensitive (M) dan longwavelength-sensitive cone. Ke-3 varian opsin tersebut terdapat pada semua sel
cones. Namun secara garis besar terdapat tiga jenis cones dimana jenisnya
tergantung pada jenis opsin yang dominan, yang menyebabkan sel ini sensitive
terhadap spectrum warna yang berbeda- beda, sebagaimana terlihat pada diagram
dibawah. Cone biru mengandung banyak blue-sensitive opsin, yang mudah
tereksitasi dengan panjang gelombang sekitar 420 nm, cone hijau dengan panjang
gelombang sekitar 530 nm, dan merah dengan panjang gelombang 560 nm.6
Trivarian colour vision
Gambar
21. Variasi
sensitivitas
warna
pada
cones
Gambar 22.A. retina sentralis didalam N. Optik
sel
25
I.
VASKULARISASI RETINA
Pembuluh darah retina berasal dari dua sumber, yaitu kapiler koroid dan
arteri dan vena sentralis. Kapiler koroid menyuplai 1/3 bagian luar termasuk sel
rod dan cone, RPE dan lapisan inti luar. Sedangkan arteri dan vena retina sentralis
menyuplai 2/3 bagian dalam sampai dengan tepi dalam lapisan inti dalam. Arteri
retina sentralis merupakan cabang pertama arteri oftalmika dengan diameter 0,3
mm dan berjalan menuju lapisan dura dari saraf optik dan memasuki bagian
inferior dan medial saraf optik sekitar 12 mm di posterior bola mata. Arteri retina
sentralis terbagi menjadi cabang superior dan inferior.
Setelah beberapa
millimeter, cabang ini terbagi menjadi cabang superior dan inferior nasal dan
temporal. Cabang dari arteri dan vena retina sentralis muncul dari bagian tengah
diskus optikus, biasanya kearah nasal. Tidak terdapat overlap dan anastomosis
pada semua pembuluh darah di semua kuadran. Cabang nasal berjalan ke ora
serrata. Sementara cabang temporal melengkung diatas dan didaerah fovea
sentralis.3
Sama halnya dengan pembuluh darah di seluruh tubuh, jika terjadi kondisi
hipoksia pada pembuluh darah retina, maka endotel dari pembuluh darah retina
akan melepaskan faktor- faktor angiogenesis seperti vascular endothelial growth
factors (VEGF), basic Fibroblast Growth Factor (bFGF), insulin-like Growth
Factor-I (IGF-I) dan berbagai nukleosida seperti adenosine (2,6,7)
AUTOREGULASI RETINA
26
Retinal blood flow pada mata yang normal selalu normal meskipun terdapat
fluktuasi pada tekanan darah sistemik maupun tekanan intraokuler. Dibandingkan
dengan sirkulasi koroid, (high-flow, variable-rate system) sirkulasi retina adalah
lower-flow, constant-rate system yang mensuplai agen-agen metabolik aktif.
Sistem saraf autonom berperan pada regulasi pembuluh darah koroid dan
retrobulbar, tetapi berakhir pada lamina kribrosa. Meskipun pembuluh darah
retina dan nervus optik memiliki reseptor -adrenergik, -adrenergik dan
kolinergik, tetapi peranan reseptor ini terhadap vaskularisasinya belum diketahui
dengan pasti. Retinal blood flow harus terjadi dalam vascular microenvironment
(autoregulation).Pada sirkulasi retina dan nervus optic, control sistemik hanya
merupakan pengaruh minor, sementara
factor local (misalnya nitrit oxide,
prostaglandin, andotelin dan system rennin-angiotensin) merupakan regulasi yang
lebih dominan.10
Pembuluh-pembuluh darah retina sangat dipengaruhi oleh sel-sel endotel,
yang melepaskan molekul vasoaktif. Molekul vasoaktif yang terpenting adalah
nitric oxide yang memicu vasodilatasi dan endothelin-1 yang memicu
vasokonstriksi. Mekanisme autoregulasi sebagian besar dikendalikan oleh
aktivitas sel neural dan glial, yang disebut neurovascular coupling, hal ini
disebabkan oleh adanya blood-retinal barrier yang menyebabkan hormon seperti
endothelin dan angiotensin hanya dapat memberi sedikit pengaruh pada
intraretina, hal ini tidak berlaku pada pembuluh darah koroid yang berfenestra,
yang memungkinkan hormon dapat lolos ke perisit dan otot polos endotel.10
AUTOREGULASI TEKANAN
Perubahan pada tekanan perfusi okuler (yang didefenisikan sebagai
Tekanan arteri pembuluh darah okuler Tekanan intraokuler, atau seringkali
dihitung 2/3 dari tekanan arteri rata-rata(mean arterial pressure) Tekanan
intraokuler) sering terjadi dalam kehidupan sehari-hari, akibat stress atau latihan
fisik yang mengakibatkan peningkatan tekanan arteri rata-rata, reduksi nocturnal
tekanan arteri dan akibat variasi diurnal TIO. Jika terjadi perubahan tekanan
perfusi, konstriksi atau dilatasi vaskuler local menyebabkan resistensi vaskuler
yang secara berlawanan meningkat atau menurun, yang berarti keadaan ini
27
mempertahankan suplai nutrien tetap stabil; Hal inilah yang dikatakan sebagai
respon autoregulasi. Karena peningkatan pada tekanan arteri brachialis sejalan
dengan tekanan arteri oftalmika, maka autoregulasi tekanan retina secara primer
dimediasi oleh meningkatnya resistensi vaskuler retina.10
AUTOREGULASI METABOLIK
Kemampuan untuk meningkatkan perfusi sebagai respon terhadap
perubahan kebutuhan jaringan, dianggap sebagai autoregulasi metabolic, yang
juga terlihat pada retina. Ketika flicker memberikan stimulasi yang meningkatkan
kebutuhan matabolik retina pada kucing atau monyet, terlihat adanya peningkatan
aliran darah retina, dengan vasodilatasi local yang dimediasi oleh pelepasan
nitrous oxide. Sebuah penelitian memperlihatkan adanya peningkatan perfusi
retina pada adaptasi gelap.
RESPON TERHADAP OKSIGEN DAN KARBON DIOKSIDA
Beberapa penelitian, menyebutkan bahwa aliran darah retina meningkat
dan menurun berlawanan dengan proporsi kandungan oksigen arteri. Pada pasien
diabetes, respon ini berkurang akibat peningkatan kadar glukosa darah. Pada
pasien sehat, sirkulasi retina merupakan cerminan sirkulasi serebral dalam hal
kandungan oksigen arteri. Faktor yang menentukan jaringan otot polos
serebrovaskuler adalah PCO2 arteri.10
Sebuah penelitian mengungkapkan manfaat penggunaan ACE inhibitor
dalam mengurangi progresifitas Retinopati Diabetik. Hal ini didasarkan pada
hipotesis bahwa terdapat sistem Renin Angiotensin yang independen pada mata.
Komponen- komponen mayor seperti reseptor angiotensin I dan II ditemukan
pada jaringan bola mata. Terdapat pula bukti bahwa ACE diproduksi oleh sel- sel
endotel dan memiliki pengaruh buruk pada aliran darah retina dan struktur
vaskuler, dan independen terhadap perubahan tekanan darah sistemik. Aktivasi
reseptor angiotensin tipe 1 pada endotel vaskuler terbukti memberi kontribusi
dalam perkembangan abnormalitas mikrovaskuler.
C. ANATOMI CORPUS VITREUS
28
C.1.Anatomi dari korpus vitreus matur
Korpus vitreus adalah suatu struktur tidak berwarna, merupakan gel transparan
yang mengisi suatu kavitas yang disebut kavitas vitreus. Korpus vitreus
mempunyai bentuk hampir spheris, kecuali bagian anterior yang mempunyai
bentuk konkaf karena adanya lensa kristalina. Korpus vitreus merupakan gel
transparan, tapi transparannya tidak homogenous. Korpus vitreus dibagi dalam
dua bagian yaitu Bagian paling luar dari korpus vitreus (atau Hyaloid), disebut
kortex yang dibagi dalam kortex anterior dan kortex posterior dan bagian dalam
yang disebut nukleus.1,4,10
Kortex vitreus
Kortex vitreus berbatasan dengan retina pada bagian posterior dan
mempunyai Densitas fibril kolagen lebih besar pada bagian perifer. Kondensasi
dari fibril kolagen ini akan membentuk suatu membrane anatomik palsu yang
disebut membrane hyaloids anterior (terletak pada anterior dari ora serrata) dan
membrane hyaloids posterior (terletak pada bagian posterior dari ora serrata).
Pada daerah antara vitreus anterior dan kapsul lensa posterior terdapat suatu
daerah yang disebut Bergers space atau disebut juga ruang retrolental erggelet.
Perlekatan kuat antara membrane hyaloid anterior dengan kapsula lensa posterior
membentuk suatu ligament yang disebut Weigerts ligament atau juga dikenal
sebagai Eggers line (hyaloideo-capsular ligament). Suatu ruangan didaerah
prepapilary yang terdapat pada bagian posterior korpus vitreus,dekat permukaan
diskus optik disebut Mortegiani space.1,10
Suatu bagian dari vitreous sekitar 2 sampai 3 mm anterior dari ora serrata,
dimana tempat ini merupakan tempat perlekatan paling kuat dari vitreus dan
memiliki ketebalan bebarapa millimeter. Daerah ini disebut Vitreus base. Vitreous
base ini juga disusun oleh fibril kolagen yang padat.1,10
Korpus Vitreus utama (Nukleus)
Nukleus merupakan bagian dari korpus vitreus yang kepadatan densitasnya
kurang (tidak sepadat kortex) sehingga membentuk struktur gel yang disebut
sebagai true biological gel. Hyaloid canal yang berjalan dari discus optic (area
Martegiani) ke posterior pole dari lensa dapat dilihat pada nukleus korpus
vitreus . Disekitar area Martegiani, lebar kanal sekitar 1-2 mm dan diarea fossa
29
patellaris yaitu sekitar 4-5 mm. Pada fetus dibelakang dari Cloquets canal
berjalan arteri hyaloids, dan arteri ini akan menghilang 6 minggu sebelum lahir
dan hyaloids canal terisi oleh cairan.1
Neurovascularisasi
Korpus vitreus tidak memiliki pembuluh darah dan serabut saraf, sehingga
meskipun pathogen telah berlangsung multipel, tidak akan mengganggu untuk
waktu yang relatif lama sebelum akhirnya muncul suatu respon immune dari
struktur didekatnya.11
Gambar 5 anatomi korpus vitreus11
30
Gambar 6 Skematik korpus vitreus bagian anterior
Keterangan gambar :
OP = Orbiculo Posterior capsular fibers, OS = Ora Serata,
OA =
Orbiculo-anterior capsular fibers, CP = Cilio-posterior capsular fibers, CA
= Cilio-equatorial capsular fibers, V = Vitreus, W = Hyaloidea - capsular
ligament of Wieger, P = Canal of Petit, H = Canal of Hannover.
C.2.Vitreoretinal interface
Vitreoretinal interface dapat diamati secara tegas dengan menggunakan
mikroskop electron, yang merupakan bagian paling luar dari kortex vitreus
(posterior hyaloid),yang terdiri dari fibril berbentuk jangkar dari vitreus dan Inner
Limitan Membrane (ILM) retina. ILM dari retina merupakan suatu struktur yang
mempunyai tebal 1 sampai 3 m, merupakan suatu membran basement yang mana
satu sisi melekat pada muller cell (footplates dari permukaan dalam retina) dan
pada sisi yang lain melekat pada korteks vitreus. Komponen yang telah
diidentifikasi pada ILM ini adalah kolagen tipe IV, kolagen tipe XVIII, laminin,
nidogen-1,agrin,perlecan dan proteoglycans.1,7
Kortex vitreus melekat dengan kuat pada ILM didaerah vitreous base, sekitar
discus optic, dibelakang lensa kristalina yaitu pada ligamentum hyloidocapsular
dari Wiegerts dan daerah sekitar foveola dengan diameter sekitar 500 m. Bila
kondisi tidak normal,pada daerah vitreoretinal interface dapat ditemukan ikatan
antara fibrils kolagen dari kortex korpus vitreus dan ILM akan melonggar.1,9
D. FISIOLOGI KORPUS VITREUS
D.1. Fungsi Korpus Vitreus
Fungsi dari korpus vitreus dapat dibagi dalam 5 group utama : 1,11
31
1. Membantu fungsi dari retina dan meningkatkan fungsi dari kavitas korpus
vitreus.
2. Sebagai barrier difusi antara segment anterior dan segment posterior bola
mata
3. Berfungsi sebagai buffer metabolic
4. Menstabilkan perjalanan cahaya (Media refrakta)
5. Konsumsi dan distribusi dari molekul oksigen
1.Membantu fungsi dari retina dan meningkatkan fungsi dari kavitas korpus
vitreus
Pada suatu kondisi normal, korpus vitreus yang intak dapat memproteksi
retina dari berbagai gangguan. Suatu korpus vitreus yang intak yang mana
mengisi bagian dalam kavitas korpus vitreus dapat menahan atau mencegah
meluasnya suatu retinal detachment. Diduga bahwa korpus vitreus dapat juga
menyerap kekuatan eksternal yang mengenai bola mata dan juga mengurangi
kerusakan mekanik terhadap bola mata, misalnya saat terjadi trauma. Korpus
vitreus yang intak juga dapat membantu lensa selama trauma terhadap kerusakan
yang lebih parah. Namun demikian mekanisme ini belum sepenuhnya benar,
karena ternyata didapatkan bahwa vitreus yang telah digerakkan melalui
vitrektomi ternyata masih berfungsi normal dan tidak terjadi retinal detachment.1
2.Sebagai barier antara segment anterior dan posterior bola mata
Pemahaman bahwa korpus vitreus adalah berbentuk gel memberikan dugaan
bahwa korpus vitreus adalah merupakan barier untuk pergerakan paling besar
antara substansi-substansi segment posterior dan anterior dari mata.1
Substansi yang berasal dari segment anterior pada mata akan sangat sukar
untuk mencapai konsentrasi tinggi pada bagian posterior mata ketika korpus
vitreus masih intak sebab difusi melalui korpus vitreus lambat dan pergerakan
alirannya terbatas oleh strukturnya yang berbentuk gel. Suatu vitreus yang intak
juga mencegah pemberian obat topikal untuk mencapai retina dan nervus optik
dengan konsentrasi yang significant. Pemberian antibiotik dari aliran darah ke
pusat korpus vitreus juga dihalangi oleh vitreus normal.
32
3.Berfungsi sebagai buffer metabolic
Pada suatu kondisi normal, ILM dan kortex posterior tidak berfungsi sebagai
barier untuk molekul yang berukuran kecil. Karena hubungan anatomi yang rapat
dari retina dan korpus siliaris, maka korpus vitreus dapat berfungsi sebagai suatu
buffer metabolik dan pada tahap tertentu dapat merupakan reservoir dari
metabolisme korpus siliaris dan terutama retina. Karena adanya blood retinal
barier, maka water soluble substance yang berlokasi pada retina dapat dengan
mudah memasuki korpus vitreus daripada ke aliran darah jika transport yang
melewati barier terbatas. 1
Substansi yang ada dalam retina atau yang juga diproduksi oleh retina dapat
berdifusi masuk ke korpus vitreus. Glukosa dan glikogen pada korpus vitreus
dapat merupakan supplement untuk metabolism retina terutama dalam kondisi
anoksia. Vitreus juga dapat berkontak dengan muller cells, dengan fungsinya
sebagai suatu buffer pada fungsi fisiologis dari muller cell, contohnya dalam
homeostasis potassium dari retina.1
4.Media refrakta
Fungsi fisiologis normal dari korpus vitreus sebagai media refrakta
disebabkan oleh sifatnya yang transparan, sehingga cahaya yang visible light
dapat sampai ke retina. Fungsi yang penting dari korpus vitreus adalah bagaimana
ia dapat menjaga transparansinya, yang secara primer dihasilkan oleh konsentrasi
rendah dari struktur makromolekul (kurang dari 0,2% berat per volume) dan
soluble protein. Transparansi dapat juga dijaga oleh kolagen spesifik atau
konfigurasi hyaluronic acid, yang dianalogikan dengan kornea dalam menjaga
transparansinya. peranannya sebagai media refrakta memberikan indeks refraktif
sekitar 1,33 yang mana hampir sama dengan indeks refraktif humor aquous.1,4
5.Konsumsi dan distribusi molekul oksigen
Konsentrasi dari ascorbat pada vitreus manusia adalah relatif terdapat dalam
konsentrasi tinggi. Pada mata dengan gel vitreus yang intak, konsentrasi rata-rata
ascorbat sekitar 2 mM. Blood levels hanya 50 sampai 60 M. Tingginya
konsentrasi ascorbat dipertahankan oleh suatu Sodium- dependent ascorbate
33
transporter (SLC23A2) pada lapisan pigmen epitel ciliaris. Peranan fisiologis dari
ascorbate pada vitreus manusia didapatkan dari investigasi dan spekulasi
experimental, tapi hasil yang didapat tidak dapat di jelaskan. Shui dan kawankawan menemukan bahwa metabolisme molekular oksigen vitreus pada suatu
ascorbate-dependent , meregulasi tekanan oksigen intraokuler.11
Sifat gel dari vitreus dengan ukuran yang luas dan berlokasi disentral dari
mata, dan dengan adanya vaskularisasi retina memberikannya oksigenasi yang
tinggi dan dengan oksigenasi yang tinggi tersebut dapat memproteksi jaringan
yang lebih sensitif terhadap oksidatif stress, seperti lensa dan trabekular
meshwork. Molekul oksigen berdifusi ke korpus vitreus dari vaskularisasi retina
akan diikat oleh ascorbat sebelum sampai ke lensa dan segment anterior. 11
Suatu hal yang tidak kalah penting, Shui dan kawan-kawan juga menemukan
bahwa
vitreus
gel
mempunyai
konsentrasi
ascorbat
lebih
tinggi
dan
mengkonsumsi oksigen lebih cepat daripada vitreus cair (misalnya pada gel
vitreus yang mengalami liquefaction atau surgical removal). Dengan demikian
mempertahankan sifat gel dari korpus vitreus adalah sangat penting. Pergerakan
transvitreal dari molekul kecil seperti oksigen tergantung pada beberapa
mekanisme seperti difusi, tekanan hidrostatik, tekanan osmotik, konveksi dan
transport aktif dari jaringan sekitar.11
Barton dan kawan-kawan baru-baru ini menunjukkan bahwa difusi molekul
kecil yang melewati vitreus gel sama dengan ketika melewati suatu cairan.
Perbedaan kritis antara pergerakan oksigen pada gel dan liquid terletak pada arus
konveksion dan cepatnya aliran dari mata. Saat vitreus dalam keadaan gel statis,
maka difusi oksigen dari retina ke gel vitreus meningkat hanya pada yang dekat
dengan jaringan retina, yang dapat ditunjukkan dengan oksigen mikroelektrode
pada penelitian expremental pada binatang percobaan. Bagaimanapun saat vitreus
mengalami pencairan maka oksigen dari pembuluh darah retina akan
didistribusikan ke mata oleh aliran cairan dan dapat terjadi oleh karena pergerakan
mata atau kepala.11
Baik oksigen maupun ascorbat akan dipakai pada reaksi dalam korpus vitreus
untuk metabolisme bola mata. Jika transport aktif ascorbat ke dalam mata konstan,
34
maka efek pencampuran oksigen akan menurunkan konsentrasi ascorbate pada
cairan vitreus,memperlambat konsumsi oksigen dan menyebabkan lebih banyak
molekul oksigen yang sampai ke lensa. Jika lensa kristalina diganti dengan suatu
intraocular lens, maka lebih banyak oksigen yang ditemukan pada trabecular
meshwork.11
Hipothesis oksigen ini konsisten dengan adanya observasi bahwa konsentrasi
ascorbate vitreus lebih rendah dan tekanan oksigen lebih tinggi pada mata dengan
pencairan vitreus atau mata yang telah diperlakukan suatu vitrektomi. Sebagai
tambahan ,bukti adanya kekonsistenan ini didapatkan pada protektif terhadap
lensa kristaline dari resiko komplikasi jangka lama post vitrektomi OAG.1
F.2.Mekanisme molekular vitreus dikaitkan dengan pertambahan umur
Korpus vitreus diduga mengalami perubahan fisiologis sepanjang hidup dari
manusia, perubahan ini memberikan perubahan yang besar terhadap fungsinya.
Suatu pola transisi dapat terlihat pada perubahan fisiologis vitreus karena umur
dan
perubahan
degeneratif
pada
vitreus
(misalnya
pada
retinitis
pigmentosa,wagner disease).1
Korpus vitreus pada postnatal normal adalah gel homogen yang mengalami
perkembangan dan konstruksi biokimia. Sebagai dasar perubahan yang terjadi
sejalan dengan pertambahan umur yaitu adanya suatu disintegrasi dari struktur
gel, yang biasa
disebut liquefaction atau synchysis, terutama pada nukleus dari
vitreus, yang mana didapatkan konsentrasi kolagen paling rendah. Liquefaction
dimulai pada awal dari kehidupan dan meningkat secara linear dengan
meningkatnya volume vitreus liquid dikaitkan dengan pertambahan umur.
Liquefaction dimulai biasanya pada daerah posterior pole dan menghasilkan
daerah atau ruangan yang disebut premacular bursa atau precortical vitreus
pocket.1
Mekanisme yang menyebabkan terjadinya liquefaction tidak dikatahui secara
pasti tapi diduga terkait dengan adanya perubahan formasi dari kolagen. Secara
35
nyata , berat molekul dari korpus vitreus akan meningkat sejalan dengan
pertambahan umur. Adapun mekanisme yang dapat menyebabkan Liquefaction
adalah: (a) adanya aktifitas enzymatic dan nonenzymatic yang melewati ikatan
kolagen fibers,(b) kerusakan kolagen fibers oleh aktifitas radikal bebas, dan (c)
penurunan densitas jaringan kolagen. Semua hal tersebut menurunkan stabilitas
dari vitreus gel.1,12
Proses liquefaction yang terjadi sejalan dengan bertambahnya umur adalah
merupakan efek kumulatif dari light exposure dan nonenzymatic glycosylation
yang dianggap sebagai penyebab terbanyak. Nonenzymatic glycosylation dapat
dihasilkan dari jaringan lain yang memiliki suatu a slow turnover dari protein
seperti pada lensa. Hyaluronic acid dan kolagen juga dapat mengalami kerusakan
oleh efek radikal bebas yang dilepaskan dari suatu photosensitizer seperti
riboflavin setelah irradiasi dengan white light.1
Dari kepustakaan ada yang berpendapat bahwa meningkatnya konsentrasi
hyaluronic acid, akan meningkatkan stabilitas dari gel, dan dianggap dengan
menurunnya hyaluronic acid akan memicu mekanisme liquefaction. Mekanisme
lain yang dianggap berperanan adalah meningkatnya soluble protein yang
dikarenakan meningkatnya kebocoran dari blood retinal barrier yang merupakan
proses fisiologis normal karena pertambahan umur atau pada suatu kondisi
pathologis seperti retinophaty diabetic.1
Dalam beberapa penelitian dengan menggunakan manipulasi
pharmakologi terhadap vitreus gel menunjukkan bahwa dengan menggunakan
hyaluronidase terjadi suatu liquefaction tanpa menginduksi terjadinya PVD
(Posterior Vitreous Detachment). Plasmin dan microplasmin juga dapat
menyebabkan terjadinya liquefaction dan menginduksi terjadinya PVD pada
beberapa pasien. 11
Dari berbagai keterangan diatas, bagaimanapun masih sulit untuk memahami
mekanisme liquefaction
korpus vitreus dengan baik, karena kurangnya
pemahaman dan kurangnya penelitian tentang hal tersebut, sangat
sulit
menggunakan binatang percobaan dalam penelitian korpus vitreus. Sebagai
alternatif, para peneliti mulai
36
mengeksplore kemungkinan menggunakan suatu substitute vitreus artificial. Dan
untuk kedepannya diharapkan substitute ini dapat digunakan untuk mempelajari
secara jelas tentang struktur dan fungsi dari vitreus gel, termasuk kemampuannya
dalam meregulasi tekanan intraocular.
BAB III
KESIMPULAN
Retina merupakan membrane yang sangat tipis dan transparan, sangat
terorganisasi dengan kemampuan untuk memulai pengolahan informasi
penglihatan sebelum ditransmisikan melalui nervus optic ke korteks visual.
Topografi retina bisa digambarkan dengan adanya macula, fovea, parafovea,
perifovea, diskus optic dan ora serata.
Pada potongan melintang dari luar ke dalam retina terdiri atas :
-
RPE dan lamina basalis
37
- Segmen luar dan dalam rods dan cones
- Membran Limitans externa
- Lapisan inti luar (nuclei fotoreceptors)
- Lapisan plexiform luar
- Lapisan inti dalam
- Lapisan plexiform dalam
- Lapisan sel ganglion
- Lapisan serat saraf (sel axon dan ganglion)
- Membran limitas interna
Fungsi retina secara keseluruhan banyak diperankan oleh Retinal Pigmen
Epithelium (RPE) :
pada siklus visual, metabolism vitamin A, blood retinal
barrier, fagositosis segmen luar fotoreseptor dan juga sebagai respon imun lokal.
Di kenal pula adanya autoregulasi pada retina.
Mata berkembang dari tiga lapisan embrional primitif yaitu ektoderm
permukaan termasuk derivatnya krista neuralis, ektoderm neuralis, dan mesoderm.
korpus vitreus sendiri berasal dari krista neuralis.
Secara anatomi korpus vitreus terbagi dalam dua bagian besar yaitu kortex
dan nukleus. kortex terdiri dari dua bagian yaitu kortex anterior dan kortex
posterior.
Korpus vitreus pada bagian anterior berbatasan dengan lensa, corpus siliaris
dan zonula, dan pada bagian posterior berbatasan dengan retina.
Korpus vitreus melekat kuat pada daerah vitreus base, kapsul posterior lensa
pada ligamentum Wiegerts,sekitar discus optik dan sekitar area fovea.
Fungsi dari korpus vitreus dibagi dalam lima group yaitu :
1. Membantu fungsi dari retina dan meningkatkan fungsi dari kavitas
korpus vitreus
2. Sebagai barrier difusi antara segment anterior dan segment posterior
bola mata
3. Berfungsi sebagai buffer metabolic
4. Menstabilkan perjalanan cahaya (Media refrakta)
5. Konsumsi dan distribusi dari molekul oksigen
Adapun mekanisme yang dapat menyebabkan Liquefaction adalah: (a)
adanya aktifitas enzymatic dan nonenzymatic yang melewati ikatan kolagen
fibers,(b) kerusakan kolagen fibers oleh aktifitas radikal bebas, dan (c) penurunan
densitas jaringan kolagen yang kesemuanya menurunkan stabilitas dari vitreus
gel.1,12
38
You might also like
- Anatomi Retina Dan VitreousDocument28 pagesAnatomi Retina Dan VitreousErlaNuraniNo ratings yet
- Anatomi RetinaDocument17 pagesAnatomi RetinaafrindaNo ratings yet
- Peranan Retina Dalam Fungsi PenglihatanDocument19 pagesPeranan Retina Dalam Fungsi Penglihatanharrisreza100% (1)
- RetinaDocument16 pagesRetinaLiyanti RinceNo ratings yet
- Retinoblastoma - ReferatDocument43 pagesRetinoblastoma - ReferatReynatta Audralia NamaraNo ratings yet
- Anatomi RetinaDocument14 pagesAnatomi RetinaJulie MckinneyNo ratings yet
- Anatomi Bola MataDocument5 pagesAnatomi Bola MataImelva GirsangNo ratings yet
- Anatomi Dan Fisiologi RetinaDocument8 pagesAnatomi Dan Fisiologi RetinaBigbig BombashNo ratings yet
- Anatomi Retina Dan VitreousDocument11 pagesAnatomi Retina Dan VitreousErlaNuraniNo ratings yet
- Bab 1Document21 pagesBab 1Hamdan Muarifin IINo ratings yet
- Anatomi RetinaDocument6 pagesAnatomi Retinadhikagugu0% (1)
- Ablasio RetinaDocument21 pagesAblasio RetinaSyarifuddin Abdul JabbarNo ratings yet
- Anatomi RetinaDocument5 pagesAnatomi RetinaAzuhra AnnisaNo ratings yet
- Ablasio RetinaDocument24 pagesAblasio RetinayuliaNo ratings yet
- RETINADocument38 pagesRETINAMuhammad RafdiNo ratings yet
- Referat Retinitis PigmentosaDocument22 pagesReferat Retinitis PigmentosaKanitia Ruzuar100% (2)
- Bab I Pendahuluan: Evoked Potentials (EP) Adalah Sinyal Bioelektrik Yang Diproduksi Oleh SistemDocument22 pagesBab I Pendahuluan: Evoked Potentials (EP) Adalah Sinyal Bioelektrik Yang Diproduksi Oleh SistemsyramadhantiNo ratings yet
- Referat Retinitis PigmentosaDocument19 pagesReferat Retinitis PigmentosaFariza AndriyawanNo ratings yet
- Refrat GWDocument42 pagesRefrat GWFrAnze TambunanNo ratings yet
- Referat Ablasio RetinaDocument22 pagesReferat Ablasio RetinaRobinder DhillonNo ratings yet
- Pemeriksaan OftalmoskopiDocument15 pagesPemeriksaan OftalmoskopiKhairani PutriNo ratings yet
- Referat Ablasio RetinaDocument16 pagesReferat Ablasio RetinaMuhammad Habib75% (4)
- Perdarahan Vitreus ReferatDocument14 pagesPerdarahan Vitreus ReferatwiwimbakNo ratings yet
- CSS Papilitis (Anat, Fisio, Defek)Document15 pagesCSS Papilitis (Anat, Fisio, Defek)alston96No ratings yet
- RetinoskopiDocument17 pagesRetinoskopiLukman Baihaqi Anar100% (1)
- Makalah Ablasio RetinaDocument17 pagesMakalah Ablasio RetinavinaNo ratings yet
- Referat RetinopatiDocument53 pagesReferat RetinopatiRahma IndahNo ratings yet
- Referat Sindrom Vogt Koyanagi HaradaDocument22 pagesReferat Sindrom Vogt Koyanagi HaradaETika Rahmi100% (1)
- Referat PapilitisDocument30 pagesReferat PapilitisAndrew Lie100% (1)
- Referat Ablasio RetinaDocument29 pagesReferat Ablasio RetinaDee GameTaemtationNo ratings yet
- Retinitis PigmentosaDocument26 pagesRetinitis PigmentosaTengku Benyamin Rusydi Jafizham33% (3)
- Oklusi Arteri Retina SentralisDocument21 pagesOklusi Arteri Retina SentralisMuhammadZaqiNo ratings yet
- Skenario 1 PBL Mata MerahDocument47 pagesSkenario 1 PBL Mata MerahSarah KemalasariNo ratings yet
- BAB I LeukokoriaDocument41 pagesBAB I LeukokoriaRefta Hermawan Laksono S67% (3)
- Anatomi RetinaDocument3 pagesAnatomi RetinaIsrianiNo ratings yet
- Referat Ablasio Retina WordDocument25 pagesReferat Ablasio Retina Wordmilda yantiNo ratings yet
- Tugas Pemeriksaan Red Refleks1Document19 pagesTugas Pemeriksaan Red Refleks1Analita Chairunissa100% (1)
- Ablasio RetinaDocument20 pagesAblasio RetinaIrfan Adi Saputra100% (3)
- Materi RetinoblastomaDocument30 pagesMateri RetinoblastomaiputusdmNo ratings yet
- RetinoblastomaDocument32 pagesRetinoblastomaMonissa ArrianiNo ratings yet
- Referat Ablasio RetinaDocument25 pagesReferat Ablasio RetinaNor Ubudiah Seti0% (1)
- Referet Amd MataDocument39 pagesReferet Amd MatapertyNo ratings yet
- REFERAT Retinopati Serosa Sentral - Dr. Dr. Nofri Suriadi, SP.MDocument21 pagesREFERAT Retinopati Serosa Sentral - Dr. Dr. Nofri Suriadi, SP.Msaleh nurazhariNo ratings yet
- RetinoskopiDocument17 pagesRetinoskopiEpifanus Arie Tanoto100% (2)
- Referat Ablasio RetinaDocument24 pagesReferat Ablasio Retinasupit1No ratings yet
- Ablasio Retina SeniorDocument31 pagesAblasio Retina SeniorHana kimmyNo ratings yet
- Anatomi RetinaDocument3 pagesAnatomi RetinaHaniaqmarNo ratings yet
- Askep Rinitis PigmentosaDocument25 pagesAskep Rinitis PigmentosaIndra SaputraNo ratings yet
- Referat AvitaminosisDocument31 pagesReferat AvitaminosisJaclinNo ratings yet
- Referat CMEDocument16 pagesReferat CMECindikia Ayu SholehaniNo ratings yet
- Referat ROP RevisiDocument31 pagesReferat ROP RevisiRatna Kurnia NingsihNo ratings yet
- Retrolental FibroplasiaDocument18 pagesRetrolental FibroplasiamiraNo ratings yet
- Bab 1Document15 pagesBab 1harzalinaNo ratings yet
- Soal OsceDocument1 pageSoal Osceriskiamanda13No ratings yet
- CVDocument2 pagesCVriskiamanda13No ratings yet
- OsceDocument15 pagesOsceriskiamanda13No ratings yet
- SURAT PengakuanDocument8 pagesSURAT Pengakuanriskiamanda13No ratings yet