You might also like
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5794)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (895)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (588)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (400)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2259)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (74)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (345)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (121)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- 1 BSC Iriigation Engineering 2018 19 Std1Document70 pages1 BSC Iriigation Engineering 2018 19 Std1Kwasi BempongNo ratings yet
- Marine Advisory 03-22 LRITDocument2 pagesMarine Advisory 03-22 LRITNikos StratisNo ratings yet
- Biology 2 Chapter 4Document61 pagesBiology 2 Chapter 4Malas Nak TaipNo ratings yet
- February 2019Document4 pagesFebruary 2019sagar manghwaniNo ratings yet
- Select Event Venue and SiteDocument11 pagesSelect Event Venue and SiteLloyd Arnold Catabona100% (1)
- Palf PDFDocument16 pagesPalf PDFKamal Nadh TammaNo ratings yet
- Birla Institute of Management and Technology (Bimtech) : M.A.C CosmeticsDocument9 pagesBirla Institute of Management and Technology (Bimtech) : M.A.C CosmeticsShubhda SharmaNo ratings yet
- Creamy and Thick Mushroom Soup: IngredientsDocument8 pagesCreamy and Thick Mushroom Soup: IngredientsSheila Mae AramanNo ratings yet
- ResearchDocument13 pagesResearchCHOYSON RIVERALNo ratings yet
- Subquery ProblemDocument9 pagesSubquery ProblemAbhi RamNo ratings yet
- Userguide SW-MC V2 2015-W45 EN S034308Document131 pagesUserguide SW-MC V2 2015-W45 EN S034308ReneNo ratings yet
- B65a RRH2x40-4R UHGC SPDocument71 pagesB65a RRH2x40-4R UHGC SPNicolás RuedaNo ratings yet
- EDAH EnglishDocument2 pagesEDAH EnglishMaría SanchoNo ratings yet
- Edgie A. Tenerife BSHM 1108: Page 1 of 4Document4 pagesEdgie A. Tenerife BSHM 1108: Page 1 of 4Edgie TenerifeNo ratings yet
- Cough PDFDocument3 pagesCough PDFKASIA SyNo ratings yet
- TFALL CaseStudy-Chandni+Chopra 072020+Document5 pagesTFALL CaseStudy-Chandni+Chopra 072020+Luis Gustavo Heredia VasquezNo ratings yet
- Analyzing Activity and Injury: Lessons Learned From The Acute:Chronic Workload RatioDocument12 pagesAnalyzing Activity and Injury: Lessons Learned From The Acute:Chronic Workload RatioLukas ArenasNo ratings yet
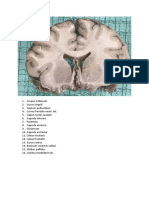
- Brain Slides SEMINAR 1 - 220606 - 142811 - 220606 - 223805Document32 pagesBrain Slides SEMINAR 1 - 220606 - 142811 - 220606 - 223805pang pangNo ratings yet
- Toaz - Info Fermentation of Carrot Juice Wheat Flour Gram Flour Etc PRDocument17 pagesToaz - Info Fermentation of Carrot Juice Wheat Flour Gram Flour Etc PRBhumika SahuNo ratings yet
- Posi LokDocument24 pagesPosi LokMarcel Baque100% (1)
- Revised List of Maharashtra HospitalsDocument16 pagesRevised List of Maharashtra Hospitalsdummy data100% (1)
- Factory OverheadDocument2 pagesFactory OverheadKeanna Denise GonzalesNo ratings yet
- Sports MedicineDocument2 pagesSports MedicineShelby HooklynNo ratings yet
- Cheap TBE Inverter TeardownsDocument33 pagesCheap TBE Inverter TeardownsWar Linux92% (12)
- High School Students' Attributions About Success and Failure in Physics.Document6 pagesHigh School Students' Attributions About Success and Failure in Physics.Zeynep Tuğba KahyaoğluNo ratings yet
- Fomula Spreadsheet (WACC and NPV)Document7 pagesFomula Spreadsheet (WACC and NPV)vaishusonu90No ratings yet
- PSB 3441 CH 1 HallucinogensDocument2 pagesPSB 3441 CH 1 HallucinogensAnonymous lm3GIU45No ratings yet
- The Chemical Composition and Organoleptic Attributes of Lesser-Known Vegetables As Consumed in Njikoka Local Government Area, Anambra State, NigeriaDocument4 pagesThe Chemical Composition and Organoleptic Attributes of Lesser-Known Vegetables As Consumed in Njikoka Local Government Area, Anambra State, NigeriaEmri CynthiaNo ratings yet
- Laws and Regulation Related To FoodDocument33 pagesLaws and Regulation Related To FoodDr. Satish JangraNo ratings yet
- Ransport and Installation Instructions CTX 510 EcoDocument32 pagesRansport and Installation Instructions CTX 510 EcoMiliano FilhoNo ratings yet