You might also like
- Embriologia Del Sistema Nervioso DR Carlos Alejandro Gonzales Medina UNMSMDocument83 pagesEmbriologia Del Sistema Nervioso DR Carlos Alejandro Gonzales Medina UNMSMCarlos GonzalesNo ratings yet
- Embriologia Power Point Ciclo OvaricoDocument22 pagesEmbriologia Power Point Ciclo OvaricoLivanessa Ubri GonzalezNo ratings yet
- Embarazo MúltipleDocument7 pagesEmbarazo MúltipleSergio BurgoaNo ratings yet
- Embarazo MúltipleDocument9 pagesEmbarazo Múltipleximena vazartNo ratings yet
- Tercera Semana Embriologia HumanaDocument33 pagesTercera Semana Embriologia HumanaJhonatan Israel Freire Liquín100% (3)
- Desarrollo de Cavidades CorporalesDocument2 pagesDesarrollo de Cavidades CorporalesArantza CuenNo ratings yet
- AnencefaliaDocument3 pagesAnencefaliaKasamivega100% (1)
- Práctica de embriología: Placenta y anexosDocument8 pagesPráctica de embriología: Placenta y anexosJohn Romero CevallosNo ratings yet
- Tercera y Octava Semana de Desarrollo EmbrionarioDocument6 pagesTercera y Octava Semana de Desarrollo EmbrionarioXiomara LuengoNo ratings yet
- Embriologia Piel Cabeza y CuelloDocument45 pagesEmbriologia Piel Cabeza y CuelloGabriel Alejandro FloresNo ratings yet
- Embriologia Del Sistema RespiratorioDocument44 pagesEmbriologia Del Sistema Respiratorioluisortega93No ratings yet
- Desarrollo Embriológico Aparato RespiratorioDocument24 pagesDesarrollo Embriológico Aparato Respiratoriojose eduardo bello ortiz100% (2)
- HISTOLOGÍADocument55 pagesHISTOLOGÍAConstanza PalavecinoNo ratings yet
- Morfogenesis de La PlacentaDocument17 pagesMorfogenesis de La PlacentajjgenconNo ratings yet
- Sindrome de PatauDocument17 pagesSindrome de PatauKrisnny MeloNo ratings yet
- Aparato Reproductor FemeninoDocument14 pagesAparato Reproductor FemeninoAngeline Cerezo Escobar100% (1)
- Embriologia Sistema CardiovascularDocument28 pagesEmbriologia Sistema CardiovascularJuanita SalasNo ratings yet
- Tema 13. Cavidades Corporales y SerosasDocument8 pagesTema 13. Cavidades Corporales y SerosasVibe NationNo ratings yet
- Aparato Reproductor FemeninoDocument32 pagesAparato Reproductor FemeninoPaula CastañedaNo ratings yet
- Sistema Cardiovascular. EMBRIOLOGIA PDFDocument10 pagesSistema Cardiovascular. EMBRIOLOGIA PDFDavid De La RosaNo ratings yet
- Desarrollo Sistema Nervioso Central S5Document48 pagesDesarrollo Sistema Nervioso Central S5LeonardoPortillaCubasNo ratings yet
- Tejido NerviosoDocument22 pagesTejido NerviosoJhon Jairo Peña GomezNo ratings yet
- Embriologia Cavidades CorporalesDocument45 pagesEmbriologia Cavidades CorporalesAbel VergaraNo ratings yet
- Intestino Medio y PosteriorDocument78 pagesIntestino Medio y PosteriorDayma SantamariaNo ratings yet
- Desarrollo Del Sistema Cardiovascular y Sus AlteracionesDocument101 pagesDesarrollo Del Sistema Cardiovascular y Sus AlteracionesNicole SalamancaNo ratings yet
- Embriologia Aparato RespiratorioDocument40 pagesEmbriologia Aparato RespiratorioByron Manuel Chicas MendezNo ratings yet
- Embriologia Sistema UrinarioDocument6 pagesEmbriologia Sistema UrinarioLizy FloresNo ratings yet
- Fecundacinsegmentacineimplantacin 121206152556 Phpapp02Document55 pagesFecundacinsegmentacineimplantacin 121206152556 Phpapp02María Esperanza Carrizo GarcíaNo ratings yet
- GUIA DE EMBRIOLOGIA C ContestadasDocument3 pagesGUIA DE EMBRIOLOGIA C Contestadasarlet mvNo ratings yet
- Glandula TimoDocument4 pagesGlandula TimoYARINo ratings yet
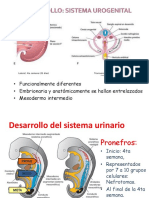
- Desarrollo Embriologico UrogenitalDocument23 pagesDesarrollo Embriologico UrogenitalClaudia Mercedes Alonso CuevasNo ratings yet
- Tromboembolia PulmonarDocument24 pagesTromboembolia PulmonarAnnia Valdivia100% (1)
- UrogenitalDocument40 pagesUrogenitalElianneNo ratings yet
- Embriologia de LagmanDocument5 pagesEmbriologia de LagmanMaria MoraNo ratings yet
- Desarrollo Embriológico Del Sistema CardiovascularDocument68 pagesDesarrollo Embriológico Del Sistema CardiovascularYoselin FontNo ratings yet
- Embriologia Del Aparato DigestivoDocument90 pagesEmbriologia Del Aparato DigestivoVicoZgNo ratings yet
- Origen y Desarrollo Del EmbarazoDocument35 pagesOrigen y Desarrollo Del EmbarazoXavier DavilaNo ratings yet
- Capitulo 55 FisioDocument2 pagesCapitulo 55 FisioVickyFingadoNo ratings yet
- Examen-Sis EsqueleticoDocument3 pagesExamen-Sis EsqueleticoLuigiZeladaNeyra100% (1)
- Prosencefalo BLDocument13 pagesProsencefalo BLbrayan_dwNo ratings yet
- ProtuberanciaDocument7 pagesProtuberanciaSteven JiménezNo ratings yet
- Cambios del embrión a través del desarrollo en la sexta semanaDocument52 pagesCambios del embrión a través del desarrollo en la sexta semanaGloria Patricia Limon BeilesNo ratings yet
- Moore Embriologia - CAPITULO UNO: Introducción Al Ser Humano en DesarrolloDocument3 pagesMoore Embriologia - CAPITULO UNO: Introducción Al Ser Humano en DesarrolloSilvia100% (1)
- Embriología de Faringe y LaringeDocument23 pagesEmbriología de Faringe y LaringeChristian Galvan67% (3)
- Embriología de Aparato Reproductor FemeninoDocument22 pagesEmbriología de Aparato Reproductor FemeninoAle Monteros100% (2)
- Semiologia de La Mama y ToraxDocument16 pagesSemiologia de La Mama y ToraxJenyfer Laurel VelasquezNo ratings yet
- Embriología 6Document25 pagesEmbriología 6Nicole DíazNo ratings yet
- Embriologia Genitales Femeninos URPDocument76 pagesEmbriologia Genitales Femeninos URPEnedina Sanchez Campos100% (2)
- Embriología - Sistema CardiovascularDocument16 pagesEmbriología - Sistema Cardiovascularjohan manuel candelario de la rosaNo ratings yet
- SINDROME DE TURNER y KlinDocument21 pagesSINDROME DE TURNER y KlinAlejandro SoteloNo ratings yet
- Anomalias CongenitasDocument37 pagesAnomalias CongenitasLina Maria Casanova GuerreroNo ratings yet
- Caso Clinico Sistema TegumentarioDocument3 pagesCaso Clinico Sistema TegumentarioYajaira GonzalesNo ratings yet
- Síndrome de WeberDocument5 pagesSíndrome de WeberHenry Brando Mallma CastroNo ratings yet
- CARDIOPATIA CONGENITA CianoticaDocument39 pagesCARDIOPATIA CONGENITA CianoticaJahirAleaxanderJibajaCedillo100% (1)
- Crecimiento y Desarrollo Embrionario y FetalDocument43 pagesCrecimiento y Desarrollo Embrionario y FetalJimmy Hoffman MarleyNo ratings yet
- Embriología Del Aparato Reproductor FemeninoDocument9 pagesEmbriología Del Aparato Reproductor FemeninoAlberty RodriguezNo ratings yet
- Placenta y Membranas FetalesssDocument47 pagesPlacenta y Membranas FetalesssStone Loqito100% (2)
- Semana 11 Práctica EmbriologíaDocument9 pagesSemana 11 Práctica EmbriologíaAna Quispe IbericoNo ratings yet
- Embriologia de EsqueletoDocument7 pagesEmbriologia de EsqueletoViviana AragonNo ratings yet
- Fisiopatología de La FiebreDocument42 pagesFisiopatología de La FiebreYuddy Equiza Barroso100% (3)
- Métodos de Diagnóstico en OrtodonciaDocument4 pagesMétodos de Diagnóstico en OrtodonciaAle BenitesNo ratings yet
- Estela AltalefDocument79 pagesEstela AltalefPatricia VelazquezNo ratings yet
- Mem-IESS-RPISSFDocument13 pagesMem-IESS-RPISSFFrancisco CarreraNo ratings yet
- Convocatoria de Posgrado 2020 21 1Document24 pagesConvocatoria de Posgrado 2020 21 1diego martinezNo ratings yet
- Fisiopatología Ganglios BasalesDocument25 pagesFisiopatología Ganglios BasalesCristopher PardoNo ratings yet
- Método JsiDocument1 pageMétodo JsiEstefanía CorcNo ratings yet
- Problemas de Estadística1Document25 pagesProblemas de Estadística1Vivis Gallegos67% (3)
- Sanación PránicaDocument2 pagesSanación PránicaMarta JorqueraNo ratings yet
- Taller Intoxicacion Alimentaria RAP2 - 03Document2 pagesTaller Intoxicacion Alimentaria RAP2 - 03Luke DiiazNo ratings yet
- DeCero A Ceto FRDocument168 pagesDeCero A Ceto FRluis gonzalitos0% (1)
- Libro de LogisticaDocument67 pagesLibro de LogisticaSullón Vargas CarlosNo ratings yet
- 4.codigo CerebroDocument17 pages4.codigo Cerebrojuan manuelNo ratings yet
- Guía para Inspección General de SSTDocument1 pageGuía para Inspección General de SSTJohnny QuenallataNo ratings yet
- Enfermedades OcupacionalesDocument8 pagesEnfermedades OcupacionalesJean Piere Macedo MezaNo ratings yet
- Santiago Henao VillegasDocument17 pagesSantiago Henao Villegasapi-228433468No ratings yet
- Dermatología CTO 3.0 PDFDocument99 pagesDermatología CTO 3.0 PDFDaniel Alvarado HernandezzNo ratings yet
- Diario 254Document39 pagesDiario 254brunoencubaNo ratings yet
- 2010-Hemorragias Disfuncionales e Hiperplasia EndometrialDocument49 pages2010-Hemorragias Disfuncionales e Hiperplasia EndometrialSusy OcampoNo ratings yet
- Clamp PDFDocument16 pagesClamp PDFAna Fernanda Hernandez SaldivarNo ratings yet
- Informe Tecnico Sobre Patologias en La ConstruccioDocument10 pagesInforme Tecnico Sobre Patologias en La ConstruccioJuan Carlos Cruz RiosNo ratings yet
- Chicaiza - Sumplementos de Calcio en GestantesDocument11 pagesChicaiza - Sumplementos de Calcio en GestantesJuanNo ratings yet
- Sistema Endocrino. GeneralidadesDocument3 pagesSistema Endocrino. GeneralidadesAngela Isabel Olvera PerezNo ratings yet
- DietoterapiaDocument7 pagesDietoterapiaDorian RiosNo ratings yet
- Manual Implementacion Protocolo de Ergonomía Res. SRT 886/15Document88 pagesManual Implementacion Protocolo de Ergonomía Res. SRT 886/15Lorena FariasNo ratings yet
- EritroplasiaDocument3 pagesEritroplasiaznachch100% (1)
- EVALUACION MODULO I de Esterilización 2021 JulioDocument4 pagesEVALUACION MODULO I de Esterilización 2021 JulionataliaNo ratings yet
- Role Impact IBCLC SpanishDocument4 pagesRole Impact IBCLC SpanishKarol Clemente BriceñoNo ratings yet
- LA MEDICINA ALTERNATIVA o COMPLEMENTARIADocument3 pagesLA MEDICINA ALTERNATIVA o COMPLEMENTARIAErnesto RamirezNo ratings yet
- Lineamientos Práctica Profesional Administración en Salud 1 Agosto 2018Document9 pagesLineamientos Práctica Profesional Administración en Salud 1 Agosto 2018wendy moraNo ratings yet