You might also like
- Taxonomy of Vascular Plants in Botanical PDFDocument45 pagesTaxonomy of Vascular Plants in Botanical PDFpepito manalotoNo ratings yet
- Taxonomy of Vascular Plants in Botanical GardenDocument46 pagesTaxonomy of Vascular Plants in Botanical Gardenpotz fernandezNo ratings yet
- Tomi Lois OlatunjiDocument16 pagesTomi Lois OlatunjiPriyono HaryonoNo ratings yet
- 04 0380 Jagessar Antimicrobial Ns PDFDocument15 pages04 0380 Jagessar Antimicrobial Ns PDFJose Rodrigo Ardila BurgosNo ratings yet
- Uji Anti Bakteri (Rubiaceae) Sebagai Tumbuhan Berkhasiat Obat Dan Upaya Perbanyaknya PDFDocument8 pagesUji Anti Bakteri (Rubiaceae) Sebagai Tumbuhan Berkhasiat Obat Dan Upaya Perbanyaknya PDFMaksar Muhuruna LaodeNo ratings yet
- Carnivorous Plants of The Worl - James Pietropaolo & Patricia P - 237501803756878Document219 pagesCarnivorous Plants of The Worl - James Pietropaolo & Patricia P - 237501803756878rgordon2No ratings yet
- Informe CalagualaDocument28 pagesInforme Calagualaカルランサ あるとぅりとNo ratings yet
- Nama Saintifik Pokok Herba @OLERICULTUREDocument22 pagesNama Saintifik Pokok Herba @OLERICULTURENabilah Abdullah NawawiNo ratings yet
- Lamiaceae Species: Biology, Ecology and Practical UsesDocument142 pagesLamiaceae Species: Biology, Ecology and Practical UsesneverwritNo ratings yet
- TA NIC Al Nomenclat UR E: Theresa Baldos Winzil Marie Cabangal Ma. Christine Mae Mansueto Richard Kyle Lesigues Group 4Document15 pagesTA NIC Al Nomenclat UR E: Theresa Baldos Winzil Marie Cabangal Ma. Christine Mae Mansueto Richard Kyle Lesigues Group 4Mica Suzanne PajeNo ratings yet
- Fchem 02 00035Document19 pagesFchem 02 00035Nick KolevNo ratings yet
- 4 Vol. 7 Issue 1 Jan 2016 IJPSR RE 1738 Paper 4Document11 pages4 Vol. 7 Issue 1 Jan 2016 IJPSR RE 1738 Paper 4Yusuf Almalik SaputraNo ratings yet
- Snake Plant InsecticideDocument48 pagesSnake Plant InsecticideAnalyn Buenaventura Clado100% (1)
- UNAAB Course Details Anatomy, Taxonomy and Physiology of Agricultural PlantsDocument26 pagesUNAAB Course Details Anatomy, Taxonomy and Physiology of Agricultural PlantsErinoso OluwadamilareNo ratings yet
- Therapeutic Uses of Carrots: A ReviewDocument7 pagesTherapeutic Uses of Carrots: A ReviewReni WulansariNo ratings yet
- Antifungal Activity of Cassia Alata LinnDocument18 pagesAntifungal Activity of Cassia Alata Linnjason gongobNo ratings yet
- 4 Vol. 12 Issue 5 May 2021 IJPSR RE 3867Document11 pages4 Vol. 12 Issue 5 May 2021 IJPSR RE 3867VinayNo ratings yet
- Laporan Awetan BasahDocument13 pagesLaporan Awetan BasahAni NurHidayantiNo ratings yet
- Review of the Medicinal Properties of the Resurrection Plant Selaginella bryopterisDocument7 pagesReview of the Medicinal Properties of the Resurrection Plant Selaginella bryopterisrazvan rotaruNo ratings yet
- Grade 12 Research TitlesDocument48 pagesGrade 12 Research TitlesJohn Louis Olerick VerdanNo ratings yet
- Literature Review of Cnidoscolus AconitifoliusDocument4 pagesLiterature Review of Cnidoscolus Aconitifoliuseubvhsvkg100% (1)
- A Review On Insulin Plant (Costus Igneus NakDocument6 pagesA Review On Insulin Plant (Costus Igneus NakKhaleel BashaNo ratings yet
- Research Paper On Plant KingdomDocument129 pagesResearch Paper On Plant KingdomRen KouenNo ratings yet
- 2017 A Mini Review On A Caralluma TuberculataDocument14 pages2017 A Mini Review On A Caralluma TuberculataSobia NoreenNo ratings yet
- Chapitredulivre BrahmietalDocument35 pagesChapitredulivre BrahmietalkokieecokNo ratings yet
- Codiaeum variegatum (Garden Croton) guideDocument4 pagesCodiaeum variegatum (Garden Croton) guideHailey RutherfordNo ratings yet
- Plants Chemical Compounds, Pharmacological and Toxicological Activity of Brugmansia Suaveolens: A ReviewDocument15 pagesPlants Chemical Compounds, Pharmacological and Toxicological Activity of Brugmansia Suaveolens: A ReviewChristian Borja TacuriNo ratings yet
- The Medicinal Prospects of Makahiya MimoDocument6 pagesThe Medicinal Prospects of Makahiya MimoDaniel BartoloNo ratings yet
- Crop Science 1 Module 1Document12 pagesCrop Science 1 Module 1Dan Miguel GabanNo ratings yet
- Aaaa PDFDocument16 pagesAaaa PDFHary EverestNo ratings yet
- Manual TaxonomiaDocument69 pagesManual Taxonomiacleicymartins1636No ratings yet
- Makabuhay Stem-Calamansi Extract Effectiveness Against CockroachesDocument28 pagesMakabuhay Stem-Calamansi Extract Effectiveness Against CockroachesChristian Dave OrateNo ratings yet
- Phys Ali San Gulat A ScienceDocument6 pagesPhys Ali San Gulat A ScienceIftri mellaniNo ratings yet
- Ficus Minahassae, HagimitDocument11 pagesFicus Minahassae, HagimitBea Bernardo100% (3)
- Cassia AlataDocument2 pagesCassia AlataQueenie LagtaponNo ratings yet
- Kistamma Singanaboina and Venkateshwar Chinna PDFDocument10 pagesKistamma Singanaboina and Venkateshwar Chinna PDFDeependra Yadav0% (1)
- Rorl Plan-DDocument11 pagesRorl Plan-DCasseylee ElerioNo ratings yet
- PROJECTDocument20 pagesPROJECTVanshika VermaNo ratings yet
- Module 3. Grow SeedlingsDocument46 pagesModule 3. Grow SeedlingsRommel Sacramento100% (1)
- Literature Review On Lantana CamaraDocument8 pagesLiterature Review On Lantana Camaraea219sww100% (1)
- Aloe vera Compounds and Their Medicinal UsesDocument14 pagesAloe vera Compounds and Their Medicinal UsesJUAN ALEJANDRO PEREZ AGUILARNo ratings yet
- Phytochemistry and Pharmacology of Callistemon Viminalis (Myrtaceae) : A ReviewDocument11 pagesPhytochemistry and Pharmacology of Callistemon Viminalis (Myrtaceae) : A ReviewikennethseemarfxNo ratings yet
- Jonathan Cariaga Stem 1 12Document12 pagesJonathan Cariaga Stem 1 12Benjie BalalaNo ratings yet
- Stellaria Media 2Document23 pagesStellaria Media 2IfraNo ratings yet
- Study of Solanaceae PlantsDocument17 pagesStudy of Solanaceae PlantsSlesha Rawal100% (1)
- Cassia Fistula Phytochemical Screening and Different ActivitiesDocument29 pagesCassia Fistula Phytochemical Screening and Different ActivitiesAnant NayabaniyaNo ratings yet
- Litsea Review BruneiDocument8 pagesLitsea Review Bruneimadbois 420No ratings yet
- Experimental Research on Maragatas' Effectiveness for DengueDocument11 pagesExperimental Research on Maragatas' Effectiveness for DenguessucbotNo ratings yet
- Classification of Botany and Use of Plants Author Instytut Zootechniki PIBDocument173 pagesClassification of Botany and Use of Plants Author Instytut Zootechniki PIBZai AtienzaNo ratings yet
- African Mistletoes Reviewed for Medicinal UsesDocument10 pagesAfrican Mistletoes Reviewed for Medicinal UsesRiscoriaNo ratings yet
- Caesalpinia CristaDocument5 pagesCaesalpinia CristaDarmawan Said100% (1)
- Visiting Taro From A Botanical and Phytochemical PerspectiveDocument6 pagesVisiting Taro From A Botanical and Phytochemical PerspectiveArpitha Shankar BandiNo ratings yet
- The Biology of Agricultural CropsDocument29 pagesThe Biology of Agricultural Cropskaye nicolasNo ratings yet
- Essential Oil Composition and Antimicrobial Interactions of Understudied Tea Tree SpeciesDocument8 pagesEssential Oil Composition and Antimicrobial Interactions of Understudied Tea Tree SpeciesxiuhtlaltzinNo ratings yet
- Thesis On Lantana CamaraDocument7 pagesThesis On Lantana Camaragbxm8h7g100% (2)
- Flora: Stavroula Mamoucha, Nikolas Fokialakis, Nikolaos S. ChristodoulakisDocument11 pagesFlora: Stavroula Mamoucha, Nikolas Fokialakis, Nikolaos S. ChristodoulakisRachmat Wihandana AgasiNo ratings yet
- Chapter One 1.1 Background of The StudyDocument6 pagesChapter One 1.1 Background of The StudyALIYU AMINU TANKONo ratings yet
- Intimacy and GriefDocument119 pagesIntimacy and GriefRobi Goco0% (1)
- Human Genetic Advancement and The Future Destruction of Todays Concept of ManDocument12 pagesHuman Genetic Advancement and The Future Destruction of Todays Concept of ManRobi GocoNo ratings yet
- The Evolution of An Egocentric Social Network in Facebook Fan PagesDocument2 pagesThe Evolution of An Egocentric Social Network in Facebook Fan PagesRobi GocoNo ratings yet
- A Resolution For Unrequited Love in The Theological PerspectiveDocument2 pagesA Resolution For Unrequited Love in The Theological PerspectiveRobi GocoNo ratings yet
- Bracts: Protective Flower LeavesDocument10 pagesBracts: Protective Flower LeavesAmeya KannamwarNo ratings yet
- Biology Investigatory ProjectDocument19 pagesBiology Investigatory ProjectPriyanks Rout50% (2)
- Detailed Lesson Plan IN Science V: Prepared By: Checked byDocument6 pagesDetailed Lesson Plan IN Science V: Prepared By: Checked byJosefina Trazona100% (12)
- Florivory of Early Cretaceous Flowers by Functionally Diverse Insects: Implications For Early Angiosperm PollinationDocument10 pagesFlorivory of Early Cretaceous Flowers by Functionally Diverse Insects: Implications For Early Angiosperm PollinationconstanzaNo ratings yet
- Concept MapDocument3 pagesConcept Mapapi-375479875No ratings yet
- Nutmeg tree fruit guideDocument16 pagesNutmeg tree fruit guideNazneen SalamNo ratings yet
- Adobe Scan Jul 19, 2023Document6 pagesAdobe Scan Jul 19, 2023Harsimran singh BawaNo ratings yet
- Anatomy of Nerium Leaf, Ovule and Anatomy of Anther 26.10.2019Document2 pagesAnatomy of Nerium Leaf, Ovule and Anatomy of Anther 26.10.2019JAGGU BHAINo ratings yet
- wk7-8 - Distinguish How Spore-Bearing and Cone-Bearing Plants ReproduceDocument28 pageswk7-8 - Distinguish How Spore-Bearing and Cone-Bearing Plants ReproduceErnita Corpuz Raymundo100% (2)
- Science 5: Quarter 2 Week 5 Module 5Document20 pagesScience 5: Quarter 2 Week 5 Module 53tj internetNo ratings yet
- Q2 W5 ScienceDocument64 pagesQ2 W5 ScienceAngie Lea Serra-YlardeNo ratings yet
- PGS 504 Notes Part IIDocument3 pagesPGS 504 Notes Part IISejal PriyaNo ratings yet
- Nueva Plantilla Curriculum Vitae Mar FondoDocument2 pagesNueva Plantilla Curriculum Vitae Mar FondoYolenis MatosNo ratings yet
- The Various Contrivances by Which Orchids Are Fertilised by InsectsDocument356 pagesThe Various Contrivances by Which Orchids Are Fertilised by InsectsJoel J. LorenzattiNo ratings yet
- Detailed Lesson Plan in The Parts of A FlowerDocument6 pagesDetailed Lesson Plan in The Parts of A FlowerCaryll Haylar Tagavilla100% (8)
- Flower, Its Structure and TypesDocument14 pagesFlower, Its Structure and TypesAtif IrfanNo ratings yet
- PTU Question PaperDocument2 pagesPTU Question PaperAgyapal MandNo ratings yet
- Perpetuation of LifeDocument1 pagePerpetuation of LifeDenise Antoneth Plangca100% (1)
- Hybrid Seed Production TechnologyDocument55 pagesHybrid Seed Production TechnologyDanish AbrarNo ratings yet
- Chapter 23 RQDocument4 pagesChapter 23 RQMia MistypuffNo ratings yet
- Double FertilizationDocument4 pagesDouble FertilizationHemanta JenaNo ratings yet
- Identifikasi TumbuhanDocument5 pagesIdentifikasi Tumbuhandiah rahmawatiNo ratings yet
- Sporophyte of PinusDocument15 pagesSporophyte of PinusMay SarohNo ratings yet
- 02-Plant Morphology (Exercise)Document5 pages02-Plant Morphology (Exercise)varshavishuNo ratings yet
- Examples of Non-Flowering Plants: GymnospermsDocument11 pagesExamples of Non-Flowering Plants: GymnospermsRogel SoNo ratings yet
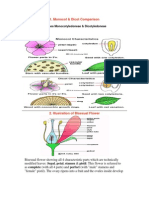
- Monocot & Dicot Comparison: Classes Monocotyledoneae & DicotyledoneaeDocument4 pagesMonocot & Dicot Comparison: Classes Monocotyledoneae & DicotyledoneaeSyan LeeNo ratings yet
- 12 Biology Imp ch2 2Document9 pages12 Biology Imp ch2 2priyanjalaproNo ratings yet
- Ch.12. Reproduction in PlantsDocument7 pagesCh.12. Reproduction in PlantsSharath ChandraNo ratings yet
- Types of Pollination: Self vs CrossDocument6 pagesTypes of Pollination: Self vs CrossKeerthi Vasan USNo ratings yet
- Parts of A Flower Crossword Puzzle Worksheet FreebieDocument7 pagesParts of A Flower Crossword Puzzle Worksheet FreebieMaite VillaltaNo ratings yet