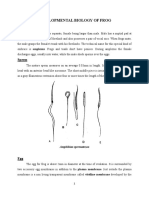
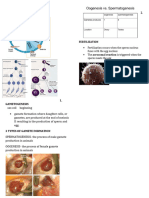
You might also like
- Camp's Zoology by the Numbers: A comprehensive study guide in outline form for advanced biology courses, including AP, IB, DE, and college courses.From EverandCamp's Zoology by the Numbers: A comprehensive study guide in outline form for advanced biology courses, including AP, IB, DE, and college courses.No ratings yet
- Frog EmbryologyDocument8 pagesFrog EmbryologyklumabanNo ratings yet
- Practical - DEVELOPMENTAL STAGES OF FROGDocument6 pagesPractical - DEVELOPMENTAL STAGES OF FROGtooba tahirNo ratings yet
- CH 47 - Animal DevelopmentDocument70 pagesCH 47 - Animal DevelopmentSofiaNo ratings yet
- Frog EmbryologyDocument3 pagesFrog Embryologyangel_soshiNo ratings yet
- Cleavage in MammalsDocument51 pagesCleavage in MammalsIrfan Azram100% (4)
- 8.introduction To EmbryologyDocument63 pages8.introduction To EmbryologyAhmed OrabyNo ratings yet
- Chapter 38 Animal DevelopmentDocument77 pagesChapter 38 Animal Developmentmaria banunaekNo ratings yet
- Animal Development 2Document7 pagesAnimal Development 2dewiNo ratings yet
- Animal - Development (Kel 1-5)Document83 pagesAnimal - Development (Kel 1-5)MelatiNo ratings yet
- Life's Greatest MiracleDocument2 pagesLife's Greatest MiracleJessa Mae Cuenco50% (4)
- Development of Frog Umanga Chapagain Read OnlyDocument9 pagesDevelopment of Frog Umanga Chapagain Read OnlySubarna PudasainiNo ratings yet
- Principles of Development: Developmental BiologyDocument86 pagesPrinciples of Development: Developmental BiologyPaolo OcampoNo ratings yet
- 7 EmbryologyDocument12 pages7 EmbryologyLansey LVSVIP16No ratings yet
- Cleavage Partitions The Zygote Into Many Smaller CellsDocument10 pagesCleavage Partitions The Zygote Into Many Smaller CellssispulieNo ratings yet
- Developmental Biology of Frog: SpermDocument21 pagesDevelopmental Biology of Frog: SpermRavindra MadurNo ratings yet
- Developmental Biology of Frog: SpermDocument21 pagesDevelopmental Biology of Frog: SpermRavindra MadurNo ratings yet
- CH-08 Notes-10.08.2021Document8 pagesCH-08 Notes-10.08.2021Chinmay SinghalNo ratings yet
- Bio 152 Lab 10 Animal Developemnt Worksheet PDFDocument18 pagesBio 152 Lab 10 Animal Developemnt Worksheet PDFHyena100% (1)
- Practical 1 GuideDocument4 pagesPractical 1 Guidea81925487No ratings yet
- GASTRULATIONDocument4 pagesGASTRULATIONProjjal SanyalNo ratings yet
- Embryology Made Ridiculously Simple Handout PDFDocument15 pagesEmbryology Made Ridiculously Simple Handout PDFAsma SaleemNo ratings yet
- Embryology Kul FKDocument65 pagesEmbryology Kul FKFahreza ArraisyNo ratings yet
- B2603 ANIMAL DEVELOPMENT - Post Fertilization EventsDocument13 pagesB2603 ANIMAL DEVELOPMENT - Post Fertilization EventssispulieNo ratings yet
- Module 5Document10 pagesModule 5Phan MhiveNo ratings yet
- Development and InheritanceDocument30 pagesDevelopment and InheritanceGeofry OdhiamboNo ratings yet
- Zebra FishDocument2 pagesZebra FishmantamadNo ratings yet
- Reproduction in AnimalsDocument3 pagesReproduction in Animalsbinu_praveenNo ratings yet
- Prenatal GrowthDocument56 pagesPrenatal GrowthnavjotsinghjassalNo ratings yet
- Human Cloning - 1Document3 pagesHuman Cloning - 1surendracrysisNo ratings yet
- Prenatal Development (Child Psychology)Document37 pagesPrenatal Development (Child Psychology)Yel BaldemorNo ratings yet
- Sexual Reproduction and Meiosis: Four Haploid CellsDocument10 pagesSexual Reproduction and Meiosis: Four Haploid CellsJohnieer Bassem MoferdNo ratings yet
- Oogenesis Data RawDocument4 pagesOogenesis Data RawMichael Angelo DejandoNo ratings yet
- Activity 6 - Animal Development 23Document10 pagesActivity 6 - Animal Development 23Via IsabelNo ratings yet
- Open Book Exam Muzzammil HussainDocument22 pagesOpen Book Exam Muzzammil HussainRIFAT RAKIBUL HASANNo ratings yet
- Exercise 2 General Features of Mammalian Zygote DevelopmentDocument8 pagesExercise 2 General Features of Mammalian Zygote DevelopmentCheska Mae ArnaizNo ratings yet
- Embryology Made Ridiculously Simple Presentation (1) Updated (1) AgainDocument100 pagesEmbryology Made Ridiculously Simple Presentation (1) Updated (1) AgainAya Sobhi100% (4)
- Class X ReproductionDocument48 pagesClass X ReproductionPagalNo ratings yet
- General Features of Chordate DevelopmentDocument66 pagesGeneral Features of Chordate DevelopmentJang WonyoungNo ratings yet
- Bio OrganismsDocument22 pagesBio OrganismsReg TbNo ratings yet
- Final Assexual - MergedDocument14 pagesFinal Assexual - Mergedpyanshy soniiNo ratings yet
- Embryological Development of The Human BrainDocument17 pagesEmbryological Development of The Human BrainErika LmNo ratings yet
- Cell Differentiation and Organogenesis 2Document34 pagesCell Differentiation and Organogenesis 2Dahal Babin100% (1)
- OAT Reading Comprehension Practice TestDocument8 pagesOAT Reading Comprehension Practice TestioatprepNo ratings yet
- 2b. Types of Egg and CleavageDocument55 pages2b. Types of Egg and CleavageManisha BishtNo ratings yet
- Only The Capacitated Spermatozoa Undergo Acrosomal ReactionDocument7 pagesOnly The Capacitated Spermatozoa Undergo Acrosomal ReactionbarbacumlaudeNo ratings yet
- Biology Lectures Finals-2Document12 pagesBiology Lectures Finals-2Jaina Alexandra MendozaNo ratings yet
- Fertilization - ImplantationDocument4 pagesFertilization - ImplantationEzeoke ChristabelNo ratings yet
- Lecture 6 - Development PDFDocument14 pagesLecture 6 - Development PDFarslanNo ratings yet
- REPRODUCTIVEDocument12 pagesREPRODUCTIVEBalolot RalphNo ratings yet
- Human Reproduction-IiDocument11 pagesHuman Reproduction-IiSuryansh SharmaNo ratings yet
- OogenesisDocument10 pagesOogenesisOm Prakash SinghNo ratings yet
- Anbt - 608: Submitted ToDocument50 pagesAnbt - 608: Submitted ToMayuriGulhaneNo ratings yet
- Human EmbryologyDocument7 pagesHuman Embryologyvinit cfuNo ratings yet
- Module 3 CytogeneticsDocument23 pagesModule 3 CytogeneticsFrances Riane SimoyNo ratings yet
- Reproduction and DevelopmentDocument121 pagesReproduction and DevelopmentAmethystNo ratings yet
- The Sea Urchin Fertilization LabDocument5 pagesThe Sea Urchin Fertilization LabAmy HollingsworthNo ratings yet
- Sofy Permana: Perkembangan Hewan DAN Ciri-Ciri Manusia Berdasarkan UsiaDocument56 pagesSofy Permana: Perkembangan Hewan DAN Ciri-Ciri Manusia Berdasarkan UsiaDhea Rafsaloka UtomoNo ratings yet
- 4th Periodical Assessment in Mathematics 7Document6 pages4th Periodical Assessment in Mathematics 7Rhonnel Manatad AlburoNo ratings yet
- K-12 Mathematics Curriculum Guide (Complete)Document94 pagesK-12 Mathematics Curriculum Guide (Complete)Rhonnel Manatad Alburo90% (166)
- Puso Making: How To Make Hanging RiceDocument1 pagePuso Making: How To Make Hanging RiceRhonnel Manatad AlburoNo ratings yet
- DepEd School Governing Council Manual - Getting StartedDocument11 pagesDepEd School Governing Council Manual - Getting StartedRhonnel Manatad Alburo100% (35)
- DepEd School Governing Council Manual - GovernanceDocument7 pagesDepEd School Governing Council Manual - GovernanceRhonnel Manatad Alburo100% (37)
- DEPED/DECS Form 137Document3 pagesDEPED/DECS Form 137Rhonnel Manatad Alburo100% (1)
- Science Investigatory ProjectDocument27 pagesScience Investigatory ProjectRhonnel Manatad Alburo90% (20)
- Strategic Intervention Material - Speed and VelocityDocument24 pagesStrategic Intervention Material - Speed and VelocityRhonnel Manatad Alburo98% (194)
- OhmDocument15 pagesOhmRhonnel Manatad Alburo88% (17)
- Mobil Delvac 1 ESP 5W-40Document3 pagesMobil Delvac 1 ESP 5W-40RachitNo ratings yet
- Toaz - Info Fermentation of Carrot Juice Wheat Flour Gram Flour Etc PRDocument17 pagesToaz - Info Fermentation of Carrot Juice Wheat Flour Gram Flour Etc PRBhumika SahuNo ratings yet
- Practice Problems Mat Bal With RXNDocument4 pagesPractice Problems Mat Bal With RXNRugi Vicente RubiNo ratings yet
- Manual Chiller Parafuso DaikinDocument76 pagesManual Chiller Parafuso Daiking3qwsf100% (1)
- Deloitte Uk Mining and Metals DecarbonizationDocument10 pagesDeloitte Uk Mining and Metals DecarbonizationfpreuscheNo ratings yet
- Generic 5S ChecklistDocument2 pagesGeneric 5S Checklistswamireddy100% (1)
- Open Cholecystectomy ReportDocument7 pagesOpen Cholecystectomy ReportjosephcloudNo ratings yet
- Test Questions For Oncologic DisordersDocument6 pagesTest Questions For Oncologic Disorderspatzie100% (1)
- LWT - Food Science and Technology: A A B ADocument6 pagesLWT - Food Science and Technology: A A B ACarlos BispoNo ratings yet
- 4th Summative Science 6Document2 pages4th Summative Science 6brian blase dumosdosNo ratings yet
- Dam Lecture 14 - Design Criteria Earth DamsDocument4 pagesDam Lecture 14 - Design Criteria Earth DamsCarolineMwitaMoseregaNo ratings yet
- Waste Sector ProjectsDocument5 pagesWaste Sector ProjectsMrcoke SeieNo ratings yet
- Social Connectedness and Role of HopelessnessDocument8 pagesSocial Connectedness and Role of HopelessnessEmman CabiilanNo ratings yet
- R633199 PDFDocument1 pageR633199 PDFChirag RohitNo ratings yet
- Preservation and Collection of Biological EvidenceDocument4 pagesPreservation and Collection of Biological EvidenceanastasiaNo ratings yet
- Capstone-ModDocument25 pagesCapstone-ModMohammad Ryyan PumbagulNo ratings yet
- Wada Defending Cannabis BanDocument18 pagesWada Defending Cannabis Banada UnknownNo ratings yet
- Complete Renold CatalogueDocument92 pagesComplete Renold CatalogueblpNo ratings yet
- Sedation and Analgesia in Critically Ill Neurologic PatientsDocument24 pagesSedation and Analgesia in Critically Ill Neurologic PatientsrazaksoedNo ratings yet
- DSM-5 Personality Disorders PDFDocument2 pagesDSM-5 Personality Disorders PDFIqbal Baryar0% (1)
- Leadership PresentationDocument10 pagesLeadership Presentationapi-404415990No ratings yet
- Artesunat Injection 2009Document4 pagesArtesunat Injection 2009Franca ImadiyiNo ratings yet
- Effect of Moisture Content On The Extraction Rate of Coffee Oil From Spent Coffee Grounds Using Norflurane As SolventDocument8 pagesEffect of Moisture Content On The Extraction Rate of Coffee Oil From Spent Coffee Grounds Using Norflurane As SolventMega MustikaningrumNo ratings yet
- Management of Developing DentitionDocument51 pagesManagement of Developing Dentitionahmed alshaariNo ratings yet
- Weekly Meal Prep GuideDocument7 pagesWeekly Meal Prep Guideandrew.johnson3112No ratings yet
- AQ-101 Arc Flash ProtectionDocument4 pagesAQ-101 Arc Flash ProtectionYvesNo ratings yet
- Water TreatmentDocument27 pagesWater TreatmentArya Singh Rathod100% (1)
- Sindh Rescue 1122 Test Sample PapersDocument12 pagesSindh Rescue 1122 Test Sample PapersMAANJONY100% (1)
- Marine Advisory 03-22 LRITDocument2 pagesMarine Advisory 03-22 LRITNikos StratisNo ratings yet
- Benefits of Silicone Coated Glass InsulatorsDocument10 pagesBenefits of Silicone Coated Glass InsulatorsMuhammad Waseem AhmadNo ratings yet