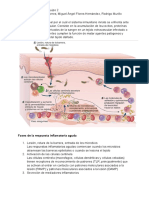
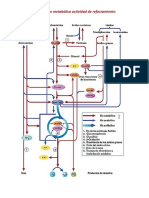
You might also like
- Digestión, Absorción y Metabolismo de Los CarbohidratosDocument51 pagesDigestión, Absorción y Metabolismo de Los CarbohidratosMeri Y Sami GómezNo ratings yet
- ProcesoDocument50 pagesProcesoPizán Estrada Luis AlbertoNo ratings yet
- Ensayo Importancia de La GlucosaDocument3 pagesEnsayo Importancia de La GlucosaEliiceoAvila50% (2)
- Tema 10 Bioquimica IIDocument4 pagesTema 10 Bioquimica IIYolanda Chaves GuerreroNo ratings yet
- Introduccion Al MetabolismoDocument17 pagesIntroduccion Al MetabolismoWilmer ParedesNo ratings yet
- GLUCOLISISDocument5 pagesGLUCOLISISLiriany CuevasNo ratings yet
- Importancia Clinica de La Membrana CelularDocument2 pagesImportancia Clinica de La Membrana CelularSARAIDY ROMERO MARLESNo ratings yet
- Ciencias Sociales Ciencias de La Vida y SaludDocument44 pagesCiencias Sociales Ciencias de La Vida y Saludcabema751038No ratings yet
- Unidad3 Carbohidratos Bioquimica ClinicaDocument10 pagesUnidad3 Carbohidratos Bioquimica ClinicaJorge MontoyaNo ratings yet
- Lipoproteínas GuíaDocument23 pagesLipoproteínas GuíaYarara YararaNo ratings yet
- Sindrome de HunterDocument18 pagesSindrome de HunterFernando Callata CardenasNo ratings yet
- Aportaciones de Otras CienciasDocument19 pagesAportaciones de Otras CienciasNelita PérezNo ratings yet
- Clase1 Alimentación y NutricionDocument38 pagesClase1 Alimentación y NutricionGabriel CorralesNo ratings yet
- Bioquimica, Ensayo Metabolismo de Carbohidratos.Document5 pagesBioquimica, Ensayo Metabolismo de Carbohidratos.DennisLandaetaNo ratings yet
- San Antonio TecomitlDocument5 pagesSan Antonio TecomitlLeonel BaronaNo ratings yet
- Glucólisis y GluconeogénesisDocument3 pagesGlucólisis y GluconeogénesisDaniel IntriagoNo ratings yet
- GluconeogenesisDocument20 pagesGluconeogenesisdan100% (2)
- Alimentación Durante El EmbarazoDocument6 pagesAlimentación Durante El EmbarazoSebastian Cristobal100% (1)
- Guia Sindrome de Hunter EspDocument13 pagesGuia Sindrome de Hunter EspYesid Yamá CuásquerNo ratings yet
- Carbohidratos ConejoDocument5 pagesCarbohidratos Conejomonica100% (1)
- Generalidades Del Tejido AdiposoDocument5 pagesGeneralidades Del Tejido AdiposoGalicia Serralde Arlet NaimNo ratings yet
- Importancia de Los ElectrolitosDocument5 pagesImportancia de Los ElectrolitosVictor Correa GarciaNo ratings yet
- 9° Economia de La SaludDocument39 pages9° Economia de La SaludMarco Aurelio Vargas Valderrama100% (1)
- InflamaciónDocument9 pagesInflamaciónIGNACIO SALVADOR FERNANDEZ GIRONNo ratings yet
- Metabolismo Es El Conjunto de Procesos Físicos y Químicos y de Reacciones A Las Que Está Sujeta Una CélulaDocument3 pagesMetabolismo Es El Conjunto de Procesos Físicos y Químicos y de Reacciones A Las Que Está Sujeta Una Célulawandres270No ratings yet
- Fisiologia Costanzo Renal-AcidobaseDocument90 pagesFisiologia Costanzo Renal-AcidobaseAbraham Serna100% (1)
- Glucolisis Descarboxilacion OxidativaDocument9 pagesGlucolisis Descarboxilacion OxidativaYoji SánchezNo ratings yet
- Alimentación, Nutrición, Salud y BienestarDocument45 pagesAlimentación, Nutrición, Salud y BienestarFatima Guadalupe Perez DiazNo ratings yet
- Biotransformacion de La EnergíaDocument82 pagesBiotransformacion de La EnergíaLia Miranda Guillen100% (3)
- Síntesis de ATP en La MitocondriaDocument2 pagesSíntesis de ATP en La MitocondriaDaniel Steven Muñoz CastilloNo ratings yet
- Anatomía Pulmonar - 19 Abril 2023 - Dr. ChávezDocument6 pagesAnatomía Pulmonar - 19 Abril 2023 - Dr. Chávezalguiensoy9512No ratings yet
- Seminario 13 - Absorción Intestinal y Secreción GástricaDocument6 pagesSeminario 13 - Absorción Intestinal y Secreción GástricaMaríaCristinaVásquezNo ratings yet
- Metabolismo de CarbohidratosDocument66 pagesMetabolismo de CarbohidratosFernanda LandetaNo ratings yet
- Mision Vuelvan CarasDocument20 pagesMision Vuelvan CarasJovir AlvarezNo ratings yet
- Rutas MetabolicasDocument30 pagesRutas Metabolicasamilcar torresNo ratings yet
- Digestion y Absorcion de Los Carbohidratos.Document12 pagesDigestion y Absorcion de Los Carbohidratos.Nayeli Elizabeth Jimenez MedinaNo ratings yet
- EndocitosisDocument6 pagesEndocitosisFhernandoRamirezNo ratings yet
- GluconeogenesisDocument37 pagesGluconeogenesisStefanny Gabriela100% (1)
- MicobacteriasDocument48 pagesMicobacteriasAbraham Kareem Madrigal RinconNo ratings yet
- 8 Metabolismo de Los LipidosDocument7 pages8 Metabolismo de Los LipidosMervin Arauz HoyosNo ratings yet
- Fosforilacion OxidativaDocument17 pagesFosforilacion OxidativaChuy Huerta Lara100% (1)
- GlucolisisDocument18 pagesGlucolisisOscar Antunez BitesNo ratings yet
- Agua y ElectrolitosDocument14 pagesAgua y ElectrolitosMayrita Ballesteros GNo ratings yet
- Ensayo de Conservacion de Micronutrientes y Macronutrientes Por Medio de Una Buena CoccionDocument7 pagesEnsayo de Conservacion de Micronutrientes y Macronutrientes Por Medio de Una Buena CoccionyeimycaraballoNo ratings yet
- Metabolismo de Las Proteínas Tarea BioquimicaDocument47 pagesMetabolismo de Las Proteínas Tarea Bioquimicacybermar2005No ratings yet
- ComensalismoDocument1 pageComensalismoedumorochitoNo ratings yet
- Derecho A La Sud en Mexico 01032023Document30 pagesDerecho A La Sud en Mexico 01032023ROGELIO ORTIZ100% (1)
- La Enfermedad de Von GierkeDocument10 pagesLa Enfermedad de Von GierkeKaren AtensioNo ratings yet
- GLUCOLISISDocument21 pagesGLUCOLISISAmeo LimaoNo ratings yet
- Metabolismo Del GLUCÓGENODocument35 pagesMetabolismo Del GLUCÓGENOErick Javier Guzman SerranoNo ratings yet
- Tema 4 Rutas Metabolicas - Carbohidratos-Lipidos-ProteinasDocument18 pagesTema 4 Rutas Metabolicas - Carbohidratos-Lipidos-Proteinasjahuer steven rueda tibaguizaNo ratings yet
- Aula 03 - El AguaDocument24 pagesAula 03 - El AguaNilson Guevara BenavidesNo ratings yet
- Afi1 U3 Atr DiomDocument4 pagesAfi1 U3 Atr Diomdira salvatoreNo ratings yet
- Modelo de Difusion y OsmosisDocument9 pagesModelo de Difusion y OsmosisMiguel Angel Ruiz GuadarramaNo ratings yet
- CARBOHIDRATOSDocument24 pagesCARBOHIDRATOSLuis Galindo100% (1)
- DISLIPIDEMIASDocument38 pagesDISLIPIDEMIASPaulina FustillosNo ratings yet
- Salud PublicaDocument7 pagesSalud Publicacri6llallaguaNo ratings yet
- 001-Digestión y Absorción de LípidosDocument27 pages001-Digestión y Absorción de LípidosGreissy Campos Cabrera100% (1)
- GlucogenólisisDocument3 pagesGlucogenólisisJosué Delgado Navas100% (1)
- Metabolismo de GlucidosDocument8 pagesMetabolismo de Glucidosmarcospicha235No ratings yet
- Tema Introducción Al MetabolismoDocument12 pagesTema Introducción Al MetabolismoMIRIAM MARTÍN SALESNo ratings yet
- Copia de Pauta Bioquímica Parcial 2Document7 pagesCopia de Pauta Bioquímica Parcial 2Nicolas MorenoNo ratings yet
- Manual Dieta Cetogenica-5Document1 pageManual Dieta Cetogenica-5Arvizu Diaz IgnacioNo ratings yet
- Piruvato DeshidrogenasaDocument2 pagesPiruvato DeshidrogenasaBraian LunaNo ratings yet
- Lectura de Metabolismo CelularDocument2 pagesLectura de Metabolismo Celulardavid.lujan.03No ratings yet
- T3 - PROCESOS ENERGETICOS EN LOS SERES VIVOS-2do Lapso - RubenrammsteinDocument8 pagesT3 - PROCESOS ENERGETICOS EN LOS SERES VIVOS-2do Lapso - RubenrammsteinrubenantonioalvarezNo ratings yet
- Not Just A Circle TraducidoDocument13 pagesNot Just A Circle TraducidovaleriaNo ratings yet
- Semana 7 Biolo TeoDocument2 pagesSemana 7 Biolo TeoKristel M. SánchezNo ratings yet
- Actividad de Reforzamiento Integracion MetabolicaDocument14 pagesActividad de Reforzamiento Integracion MetabolicaJosselyn GaynorNo ratings yet
- Respiracion Aerobica Y AnaerobicaDocument6 pagesRespiracion Aerobica Y AnaerobicaDaniela A QuezadaNo ratings yet
- 1°medio Biologia Guia 1 AgostoDocument4 pages1°medio Biologia Guia 1 AgostomaricelNo ratings yet
- Bio Qui MicaDocument7 pagesBio Qui MicaLUIS ANDRES CRUCES MUSTTONo ratings yet
- Proced I Mien ToDocument3 pagesProced I Mien ToMayraSamameBurgosNo ratings yet
- BioquímicaDocument26 pagesBioquímicaVale Ignacia Farias ReyesNo ratings yet
- rdtgjkTEMA 28Document16 pagesrdtgjkTEMA 28Arturo GálvezNo ratings yet
- 17 Condicion Corporal y Cetosis IDocument4 pages17 Condicion Corporal y Cetosis IAxoncomunicacionNo ratings yet
- Guia Metabolismo Lipidico 2022Document6 pagesGuia Metabolismo Lipidico 2022PaumelinNo ratings yet
- Elabora Modelos BidiomensionalesDocument4 pagesElabora Modelos Bidiomensionalesusuario lopezNo ratings yet
- Fotosíntesis y Respiración CelularDocument4 pagesFotosíntesis y Respiración CelularaleNo ratings yet
- Actividad 1. ¿Necesitamos Las Plantas paraDocument2 pagesActividad 1. ¿Necesitamos Las Plantas paraPoncho Baez CordovaNo ratings yet
- PASOS PREVIO de La Beta OxidacionDocument6 pagesPASOS PREVIO de La Beta OxidaciongiovannaNo ratings yet
- Quimica Premedico Medicina Integral ComunitariaDocument7 pagesQuimica Premedico Medicina Integral ComunitariaRoss MorinNo ratings yet
- Qué Es El MetabolismoDocument3 pagesQué Es El MetabolismoAbigail JimenezNo ratings yet
- Metabolitos Primarios y Secundarios de Las PlantasDocument27 pagesMetabolitos Primarios y Secundarios de Las PlantasANA GRIJALVANo ratings yet
- Apuntes - Eliminación de AmonioDocument5 pagesApuntes - Eliminación de AmonioLaura AybarNo ratings yet
- Eda 8 Act.1 - CtaDocument3 pagesEda 8 Act.1 - CtaLUCKY ғғNo ratings yet
- Hematologia Completa Y Plaquetas: MG/DLDocument2 pagesHematologia Completa Y Plaquetas: MG/DLAlan Manuel Rojas ANo ratings yet
- Relacion de La Síntesis de Acidos Grasos Con El Metabolismo de Los CarbohidratosDocument3 pagesRelacion de La Síntesis de Acidos Grasos Con El Metabolismo de Los CarbohidratosPedro Manzanilla100% (3)
- Vía de Las Pentosas Fosfato CACADocument6 pagesVía de Las Pentosas Fosfato CACACarlos CastroNo ratings yet
- Clase 4 Formulas IafDocument26 pagesClase 4 Formulas IafCayamuc BetancourtNo ratings yet