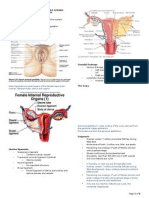
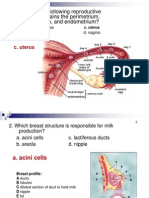
You might also like
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5783)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (890)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (399)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (587)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (265)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (72)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (344)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2219)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (119)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- Functions of PlacentaDocument8 pagesFunctions of PlacentappsavidalandNo ratings yet
- Chemical Coordination and Integration: SolutionsDocument10 pagesChemical Coordination and Integration: SolutionsIhtisham Ul HaqNo ratings yet
- Pregnancy TestDocument2 pagesPregnancy TestGianna SablanNo ratings yet
- Auctores: Clinical Medical Reviews and ReportsDocument4 pagesAuctores: Clinical Medical Reviews and ReportsMohamed AlkaliphyNo ratings yet
- Ovulation Induction Evidence Based GuidelinesDocument207 pagesOvulation Induction Evidence Based Guidelineswaleedali100% (3)
- Functions of PlacentaDocument18 pagesFunctions of PlacentaMohammad NabeelNo ratings yet
- NASBR Final Abstracts 181003 PDFDocument98 pagesNASBR Final Abstracts 181003 PDFJose Juan FloresNo ratings yet
- Endocrine System OverviewDocument23 pagesEndocrine System OverviewKimberly Joy GregorioNo ratings yet
- Luteinizing HormoneDocument8 pagesLuteinizing HormoneSaba MalikNo ratings yet
- Management Strategies For Poseidon 3 and 4Document11 pagesManagement Strategies For Poseidon 3 and 4Divisi FER MalangNo ratings yet
- Modeling Endocrine Control of The Pituitary-Ovarian Axis: Androgenic Influence and Chaotic DynamicsDocument22 pagesModeling Endocrine Control of The Pituitary-Ovarian Axis: Androgenic Influence and Chaotic DynamicsKerin ArdyNo ratings yet
- Fish Breeding, Nursing and TransportationDocument43 pagesFish Breeding, Nursing and TransportationJogis SuciNo ratings yet
- DR - Asdala TransDocument8 pagesDR - Asdala TransthorheavenNo ratings yet
- InVitroFertilization IVF KP AnalysisDocument14 pagesInVitroFertilization IVF KP AnalysisOvn Murthy100% (1)
- DR Khaled Final Exam MCQ 2012 - 2013Document89 pagesDR Khaled Final Exam MCQ 2012 - 2013ﻣﻠﻚ عيسى100% (2)
- AmenorrheaDocument148 pagesAmenorrheaAbhishek VijayakumarNo ratings yet
- B29061KDocument13 pagesB29061KMeethuanNo ratings yet
- OB Power Point Presntation 001Document50 pagesOB Power Point Presntation 001RitamariaNo ratings yet
- WEEK 1 SLM 1 Hormones in Reproductive Systems SLM 1Document12 pagesWEEK 1 SLM 1 Hormones in Reproductive Systems SLM 1yuri sabileNo ratings yet
- Carp Culture notes-II 1. Process and Factors Influencing Maturation of FishDocument19 pagesCarp Culture notes-II 1. Process and Factors Influencing Maturation of FishSabia Khan Sabia KhanNo ratings yet
- SamarthDocument11 pagesSamarthajay30No ratings yet
- Role of Serotonin in Fish Reproduction: Parvathy Prasad, Satoshi Ogawa and Ishwar S. ParharDocument9 pagesRole of Serotonin in Fish Reproduction: Parvathy Prasad, Satoshi Ogawa and Ishwar S. ParharPuri RahmaNo ratings yet
- DisxussionDocument2 pagesDisxussionGERA KAYE BUNCALANNo ratings yet
- H-046-003258-00 Total β HCG KIT (CLIA) Muti laguageDocument15 pagesH-046-003258-00 Total β HCG KIT (CLIA) Muti laguageSinari AlfatNo ratings yet
- Development and Structure of the Mammalian PlacentaDocument8 pagesDevelopment and Structure of the Mammalian PlacentaZulfi Nur Amrina RosyadaNo ratings yet
- Anabolic SteroidsDocument56 pagesAnabolic Steroidstrungkunming100% (1)
- Insert - LH.ms 11732234122.v19.enDocument4 pagesInsert - LH.ms 11732234122.v19.enyantuNo ratings yet
- CH 18 Lecture PresentationDocument127 pagesCH 18 Lecture PresentationMaski03No ratings yet
- MCQ On Menstrual CycleDocument7 pagesMCQ On Menstrual CycleKarishma Shroff100% (1)
- Sun 800 RoidRage Innovative Trends Treatment Male HypogonadismDocument19 pagesSun 800 RoidRage Innovative Trends Treatment Male HypogonadismM. Sz.No ratings yet