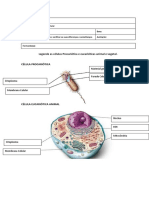
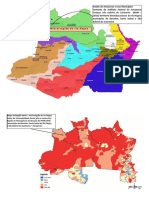
You might also like
- Relatório de Avaliação Tratamento de Efluentes - ESALQDocument40 pagesRelatório de Avaliação Tratamento de Efluentes - ESALQchicoaqueirozNo ratings yet
- TCC Treino de Força EscaladaDocument47 pagesTCC Treino de Força EscaladachicoaqueirozNo ratings yet
- Avaliação Da Disciplina Sistemas de Gestão AmbientalDocument2 pagesAvaliação Da Disciplina Sistemas de Gestão AmbientalchicoaqueirozNo ratings yet
- Gerenciamento de Resíduos (10!05!13) Versao Final PedóDocument30 pagesGerenciamento de Resíduos (10!05!13) Versao Final PedóchicoaqueirozNo ratings yet
- Chave para Restauração Florestal PDFDocument12 pagesChave para Restauração Florestal PDFchicoaqueirozNo ratings yet
- REFERÊNCIA Avefauna Santa GenebraDocument19 pagesREFERÊNCIA Avefauna Santa GenebrachicoaqueirozNo ratings yet
- A Importância Dos Corredores para A FaunaDocument10 pagesA Importância Dos Corredores para A FaunarfarneziNo ratings yet
- REFERENCIA Plasticidade RhinellaDocument32 pagesREFERENCIA Plasticidade RhinellachicoaqueirozNo ratings yet
- Aves Brasil 2014Document42 pagesAves Brasil 2014chicoaqueirozNo ratings yet
- A Prática de Mindfulness Módulo IDocument29 pagesA Prática de Mindfulness Módulo IMário Oliveira100% (1)
- Ciências Neurológicas AvaliacaoDocument4 pagesCiências Neurológicas AvaliacaoPâmela Azevedo0% (1)
- 2014 Envelhecimento Cutâneo 1Document274 pages2014 Envelhecimento Cutâneo 1Paulo Adrian Assunção da Silva100% (1)
- INEA - DZ 209.R 2pdfDocument3 pagesINEA - DZ 209.R 2pdfCarlosjr83No ratings yet
- Nematoides em HortalicasDocument196 pagesNematoides em Hortalicasanderson_rbNo ratings yet
- D6 (3 Série - Ens. Médio - L.P - BLOG Do Prof. Warles)Document15 pagesD6 (3 Série - Ens. Médio - L.P - BLOG Do Prof. Warles)Alex KayoNo ratings yet
- Ficha de TrabalhoDocument3 pagesFicha de TrabalhoGabriella 2No ratings yet
- Kit Brasil LivingDocument15 pagesKit Brasil LivingMaite ArquivoNo ratings yet
- Resumo Artigo Conceito de Bioma Coutinho 2006Document2 pagesResumo Artigo Conceito de Bioma Coutinho 2006Roberta MirandaNo ratings yet
- Adeuspsoriase PDFDocument88 pagesAdeuspsoriase PDFFernanda Amaroke100% (2)
- MAPA - Amazonas - Microrregião Do Rio Negro IDH-IVS e Outras NotasDocument8 pagesMAPA - Amazonas - Microrregião Do Rio Negro IDH-IVS e Outras NotasSouza EliasNo ratings yet
- PDF CCD2 Aula 1 Ósseas e ArticularesDocument27 pagesPDF CCD2 Aula 1 Ósseas e ArticularesJuliane Araújo LimaNo ratings yet
- Portfólio de PlantasDocument20 pagesPortfólio de PlantasEduardo LampugnaniNo ratings yet
- Mensagem A Garcia Elbert HubbardDocument8 pagesMensagem A Garcia Elbert HubbardJean CasuccioNo ratings yet
- Como Pensar e Viver Melhor - Rolf DobelliDocument340 pagesComo Pensar e Viver Melhor - Rolf DobelliAlgoritmo Fono100% (1)
- Espermograma - Análise Do SêmemDocument12 pagesEspermograma - Análise Do SêmemMarilene LopesNo ratings yet
- 1 5179326943719326206Document246 pages1 5179326943719326206Pri PerolitasNo ratings yet
- CV ExtraDocument2 pagesCV ExtraMATILDE FERREIRA MARTINSNo ratings yet
- PCC Psicologia Do Desenvolvimento e Da AprendizagemDocument11 pagesPCC Psicologia Do Desenvolvimento e Da AprendizagemJéssica Santos50% (6)
- 7 Bri, Pteri, Ginos, Angiosperma 7 EJADocument3 pages7 Bri, Pteri, Ginos, Angiosperma 7 EJAglicia joamaNo ratings yet
- A Evolução e Eu - George Gilder (Artigo)Document16 pagesA Evolução e Eu - George Gilder (Artigo)Ricardo MaddalenaNo ratings yet
- Tarzan, O Invencivel - Tarzan - Edgar Rice BurroughsDocument165 pagesTarzan, O Invencivel - Tarzan - Edgar Rice BurroughswaldimiroNo ratings yet
- Curso de Telepatia deDocument35 pagesCurso de Telepatia deClaudio Rocha Oliveira SilvaNo ratings yet
- 369785-Fosforilação OxidativaDocument4 pages369785-Fosforilação OxidativaRamires Cerqueira100% (1)
- Conservação de Aves Endemicas Ameacadas de PE LivroDocument80 pagesConservação de Aves Endemicas Ameacadas de PE Livrocarolquimica24No ratings yet
- Laboratório Municipal de Saúde Pública Do Recife Prof. Oswaldo Gonçalves de LimaDocument1 pageLaboratório Municipal de Saúde Pública Do Recife Prof. Oswaldo Gonçalves de LimaRicardo CabralNo ratings yet
- Efeito de 12 Semanas de Treinamento ResistidoDocument6 pagesEfeito de 12 Semanas de Treinamento ResistidoYk TreinamentoNo ratings yet
- Aula 1 ArrozDocument72 pagesAula 1 ArrozNelson CossaNo ratings yet
- 01 Radiobiologia RadioproteçãoDocument91 pages01 Radiobiologia Radioproteçãohugolemos2710No ratings yet
- Absorção Vitamina DDocument1 pageAbsorção Vitamina DHeide Cristina Alves DiasNo ratings yet