You might also like
- Microbial Plant Pathogens: Detection and Management in Seeds and PropagulesFrom EverandMicrobial Plant Pathogens: Detection and Management in Seeds and PropagulesNo ratings yet
- Occurrence and Detection of Sweet Potato Virus DiseaseDocument4 pagesOccurrence and Detection of Sweet Potato Virus DiseaseShailendra RajanNo ratings yet
- Moulds and Mycotoxins in Stored Maize Grains PDFDocument10 pagesMoulds and Mycotoxins in Stored Maize Grains PDFTiara Febriani ChaesarioNo ratings yet
- 1 s2.0 0168170286900080 MainDocument13 pages1 s2.0 0168170286900080 MainfalmazanNo ratings yet
- Grupo 8 - M Bovis PDFDocument13 pagesGrupo 8 - M Bovis PDFClaudia Salgado MuñozNo ratings yet
- 270 1 116 PDFDocument8 pages270 1 116 PDFJulian PinedaNo ratings yet
- Barbier Et Al 2003 SalsomaggioreDocument4 pagesBarbier Et Al 2003 SalsomaggioreennescribeNo ratings yet
- Biology of Cotton Mealybug, Phenacoccus Solenopsis (Tinsley) On Some Vegetable Crops in The Gazira State, SudanDocument4 pagesBiology of Cotton Mealybug, Phenacoccus Solenopsis (Tinsley) On Some Vegetable Crops in The Gazira State, SudanIJEAB JournalNo ratings yet
- 1 PBDocument9 pages1 PBAustine OsaweNo ratings yet
- Antifungal Activity of Eugenol Against Botrytis Cinerea: Chunmei Wang, Jie Zhang, Hao Chen, Yongjian FanDocument7 pagesAntifungal Activity of Eugenol Against Botrytis Cinerea: Chunmei Wang, Jie Zhang, Hao Chen, Yongjian FanTazyinul Qoriah AlfauziahNo ratings yet
- Bacterial and Viral Pathogens AssociatedDocument10 pagesBacterial and Viral Pathogens Associatednanochabow537No ratings yet
- Molecular Characterization of Extended-Spectrum BetaLactamase (ESBLs) Genes in Pseudomonas Aeruginosa From Pregnant Women Attending A Tertiary Health Care Centre in Makurdi, Central NigeriaDocument7 pagesMolecular Characterization of Extended-Spectrum BetaLactamase (ESBLs) Genes in Pseudomonas Aeruginosa From Pregnant Women Attending A Tertiary Health Care Centre in Makurdi, Central NigeriaJASH MATHEWNo ratings yet
- Prevalence of Mec A Gene Positive Coagulase Negative StaphyloDocument6 pagesPrevalence of Mec A Gene Positive Coagulase Negative Staphylosuresh151971No ratings yet
- 2023 Gorman High Throughput DNA Extraction PlateDocument9 pages2023 Gorman High Throughput DNA Extraction PlateCamila GamboaNo ratings yet
- Detection of Alternaria Radicina and A Dauci From Imported Carrot Seed in New ZealandDocument7 pagesDetection of Alternaria Radicina and A Dauci From Imported Carrot Seed in New ZealandPaula Ordenes DomínguezNo ratings yet
- Article ReviewDocument16 pagesArticle ReviewTasrin PrityNo ratings yet
- Comparative Comprehensive Analysis On Natural InfeDocument15 pagesComparative Comprehensive Analysis On Natural InfeThat alnetaqainNo ratings yet
- Krzysztof Treder, Włodzimierz Przewodowski, Agnieszka BarnykDocument10 pagesKrzysztof Treder, Włodzimierz Przewodowski, Agnieszka BarnykKrzysztof TrederNo ratings yet
- Emma - Pdis 06 22 1350 SCDocument3 pagesEmma - Pdis 06 22 1350 SCOlajide Emmanuel OlorunfemiNo ratings yet
- Transmission of MWMV by Aphids 2016Document6 pagesTransmission of MWMV by Aphids 2016Inma Espinar VergelNo ratings yet
- Distribution of SCMV On APPSDocument3 pagesDistribution of SCMV On APPSLilik K PutraNo ratings yet
- Extent of Microbial Contamination of Sausages Sold in Two Nigerian CitiesDocument4 pagesExtent of Microbial Contamination of Sausages Sold in Two Nigerian CitiesupyluthNo ratings yet
- 2 MycopathDocument6 pages2 MycopathPangeran Kelinci ßNo ratings yet
- ID None PDFDocument6 pagesID None PDFJamrud KhatulistiwaNo ratings yet
- EJAR - Volume 101 - Issue 3 - Pages 739-750Document12 pagesEJAR - Volume 101 - Issue 3 - Pages 739-750nailulNo ratings yet
- The Diagnostic Yield of Acid-Fast-Bacillus Smear-Positive Sputum SpecimensDocument2 pagesThe Diagnostic Yield of Acid-Fast-Bacillus Smear-Positive Sputum SpecimensNaye AbarcaNo ratings yet
- EJHM Volume 77 Issue 6 Pages 5804-5816Document13 pagesEJHM Volume 77 Issue 6 Pages 5804-5816فردوس يسNo ratings yet
- JurnalDocument5 pagesJurnalAstiNo ratings yet
- 10.1515 - ZNC 2009 5 618Document6 pages10.1515 - ZNC 2009 5 618Sona HaciliNo ratings yet
- Bacteriology of Tonsil Surface and Core in Children With Chronic Tonsillitis and Incidence of Bacteraemia During TonsillectomyDocument11 pagesBacteriology of Tonsil Surface and Core in Children With Chronic Tonsillitis and Incidence of Bacteraemia During TonsillectomyAkatsuki HendriNo ratings yet
- Technique Detection Spongospora in Soil 1983 SzeDocument2 pagesTechnique Detection Spongospora in Soil 1983 SzeDario BaronaNo ratings yet
- Prevalence of Salmonella Enteritidis Infection in Chicken Eggs Collected From Different Poultry Farms in Awka, Anambra State of NigeriaDocument11 pagesPrevalence of Salmonella Enteritidis Infection in Chicken Eggs Collected From Different Poultry Farms in Awka, Anambra State of NigeriaEditor IjrssNo ratings yet
- Monitoring The Ecology of Bacillus During Daqu In-Wageningen University and Research 291543Document10 pagesMonitoring The Ecology of Bacillus During Daqu In-Wageningen University and Research 291543Muhammad RifqiNo ratings yet
- Department of Biology Department of Microbiology College of Education Medicine College Mosul University Salah-Edin UniversityDocument7 pagesDepartment of Biology Department of Microbiology College of Education Medicine College Mosul University Salah-Edin UniversityAhmed J AlhindaweNo ratings yet
- Early Embryo Invasion As A Determinant in Pea of The Seed Transmission of Pea Seed-Borne Mosaic VirusDocument6 pagesEarly Embryo Invasion As A Determinant in Pea of The Seed Transmission of Pea Seed-Borne Mosaic VirusDenih WahyudinNo ratings yet
- Isolation and Identification of Streptomyces From Different Sample of SoilsDocument6 pagesIsolation and Identification of Streptomyces From Different Sample of SoilsMandy FisherNo ratings yet
- tmpB33A TMPDocument10 pagestmpB33A TMPFrontiersNo ratings yet
- Suay 2000 ATMB BasidyoDocument11 pagesSuay 2000 ATMB Basidyoinositol66No ratings yet
- Article 12176Document9 pagesArticle 12176Raynaldi PrabowoNo ratings yet
- Distribution of Salmonella Serovars and Antimicrobial Susceptibility of Salmonella Enteritidis From Poultry in ZimbabweDocument7 pagesDistribution of Salmonella Serovars and Antimicrobial Susceptibility of Salmonella Enteritidis From Poultry in ZimbabweDulce Amor100% (1)
- Molecular Aproach Detection of BabseieDocument11 pagesMolecular Aproach Detection of BabseieAamir muse osmanNo ratings yet
- Genetic Dissimilarity of Commensal of Carried in Different Anatomical Locations of HealthyDocument9 pagesGenetic Dissimilarity of Commensal of Carried in Different Anatomical Locations of Healthyfaithfull_802061No ratings yet
- 499-Article Text-706-1-10-20220526-MutoniDocument8 pages499-Article Text-706-1-10-20220526-MutoniChristine MutoniNo ratings yet
- Mass Production of AM Fungal Inoculum by Soil Based Pot CultureDocument5 pagesMass Production of AM Fungal Inoculum by Soil Based Pot CultureIJARBS JOURNALNo ratings yet
- TMP 1 FEFDocument7 pagesTMP 1 FEFFrontiersNo ratings yet
- Makalah Seminar Untuk PerbaikanDocument10 pagesMakalah Seminar Untuk PerbaikanasihNo ratings yet
- Jurnal BakteriologiDocument7 pagesJurnal BakteriologiEva NadilaNo ratings yet
- Anale Iasi 2012Document14 pagesAnale Iasi 2012sofia2010monicaNo ratings yet
- 9Document3 pages9kicklokickNo ratings yet
- Schoenanthus: Kassem, S. A. A. M. A. M. Abo-Tayesh and H. M. M.HeikalDocument9 pagesSchoenanthus: Kassem, S. A. A. M. A. M. Abo-Tayesh and H. M. M.HeikalBalaji MohantyNo ratings yet
- Sorho - 2014 - Identification of Viral Disease of Solanaceus in CIDocument8 pagesSorho - 2014 - Identification of Viral Disease of Solanaceus in CIDio DembeleNo ratings yet
- Otro Articulo PDFDocument9 pagesOtro Articulo PDFRomeroNo ratings yet
- International Journal of Scientific Research: MicrobiologyDocument3 pagesInternational Journal of Scientific Research: MicrobiologyPriyanka misaleNo ratings yet
- Sources of Microbial Contamination in TC LabDocument6 pagesSources of Microbial Contamination in TC LabLau Shin YeeNo ratings yet
- APS Abstracts Hawaii, USA 2011Document206 pagesAPS Abstracts Hawaii, USA 2011ggajendranNo ratings yet
- 3850 8696 1 PBDocument10 pages3850 8696 1 PBYesiNo ratings yet
- Pinot Et Al-2011-Journal of Applied MicrobiologyDocument9 pagesPinot Et Al-2011-Journal of Applied Microbiologydr Alex stanNo ratings yet
- EAJBSC-Volume 15-Issue 1 - Page 455-473Document20 pagesEAJBSC-Volume 15-Issue 1 - Page 455-473nailulNo ratings yet
- Eivaziankary2018 PDFDocument8 pagesEivaziankary2018 PDFGiribabu PalaniappanNo ratings yet
- Soberhea DermatisDocument5 pagesSoberhea DermatisRisma PertiwiNo ratings yet
- SugaNate 160NCDocument5 pagesSugaNate 160NCmndmattNo ratings yet
- JavaScript ArraysDocument5 pagesJavaScript Arraysursu_padure_scrNo ratings yet
- Журнал JournalistDocument36 pagesЖурнал JournalistSofia SorochinskaiaNo ratings yet
- Do You Have Esp FinalDocument20 pagesDo You Have Esp FinalVrasidas Poulopoulos50% (2)
- Gerin Bagaslino - 52417512Document13 pagesGerin Bagaslino - 52417512Gerin BagaslinoNo ratings yet
- BeginnersGuide AndroidonOmapZoomDocument10 pagesBeginnersGuide AndroidonOmapZoomm13marleNo ratings yet
- OS Lab ManualDocument47 pagesOS Lab ManualNivedhithaVNo ratings yet
- Oliver vs. Philippine Savings BankDocument14 pagesOliver vs. Philippine Savings BankAmeir MuksanNo ratings yet
- Federal Register / Vol. 72, No. 108 / Wednesday, June 6, 2007 / Proposed RulesDocument1 pageFederal Register / Vol. 72, No. 108 / Wednesday, June 6, 2007 / Proposed RulesJustia.comNo ratings yet
- A Theory of Ethnic Group BoundariesDocument28 pagesA Theory of Ethnic Group BoundariesAbdellatif El BekkariNo ratings yet
- Canada Post ReportDocument202 pagesCanada Post ReportrgranatsteinNo ratings yet
- Practical Means For Energy-Based Analyses of Disproportionate Collapse PotentialDocument13 pagesPractical Means For Energy-Based Analyses of Disproportionate Collapse PotentialNhut NguyenNo ratings yet
- DBR G+24 Building ProjectDocument28 pagesDBR G+24 Building ProjectShobhit Mohta100% (1)
- MAK Halliday The Language of ScienceDocument268 pagesMAK Halliday The Language of ScienceJeneiGabriellaOfő100% (4)
- Property PalleteDocument5 pagesProperty PalletePranay PatelNo ratings yet
- What Are The Subjects Which Come in UPSC Engineering Services - Electronics - Communication - ExamDocument2 pagesWhat Are The Subjects Which Come in UPSC Engineering Services - Electronics - Communication - ExamVikas ChandraNo ratings yet
- Wa0004Document8 pagesWa0004Thabo ChuchuNo ratings yet
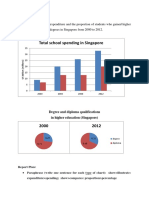
- Total School Spending in Singapore: Task 1 020219Document6 pagesTotal School Spending in Singapore: Task 1 020219viper_4554No ratings yet
- Kaizen Idea - Sheet: Suitable Chairs To Write Their Notes Faster & Easy Method While TrainingDocument1 pageKaizen Idea - Sheet: Suitable Chairs To Write Their Notes Faster & Easy Method While TrainingWahid AkramNo ratings yet
- IBRO News 2004Document8 pagesIBRO News 2004International Brain Research Organization100% (1)
- Network Management Systems 10CS834 PDFDocument113 pagesNetwork Management Systems 10CS834 PDFSarfraz AhmedNo ratings yet
- DSpace Configuration PDFDocument1 pageDSpace Configuration PDFArshad SanwalNo ratings yet
- Segmented Shaft Seal Brochure Apr 08Document4 pagesSegmented Shaft Seal Brochure Apr 08Zohaib AnserNo ratings yet
- Control Panel Manual 1v4Document52 pagesControl Panel Manual 1v4Gustavo HidalgoNo ratings yet
- 2.2 Lesson Notes FA12Document24 pages2.2 Lesson Notes FA12JAPNo ratings yet
- Organizational Behavior - Motivational Theories at Mcdonald's ReportDocument11 pagesOrganizational Behavior - Motivational Theories at Mcdonald's ReportvnbioNo ratings yet
- People in Organization: Group - 6Document8 pagesPeople in Organization: Group - 6rennys amaliaNo ratings yet
- For Purposeful and Meaningful Technology Integration in The ClassroomDocument19 pagesFor Purposeful and Meaningful Technology Integration in The ClassroomVincent Chris Ellis TumaleNo ratings yet
- Kina23744ens 002-Seisracks1Document147 pagesKina23744ens 002-Seisracks1Adrian_Condrea_4281No ratings yet
- Unit 9 Caring and Sharing Year 2 KSSR Lesson PlanDocument6 pagesUnit 9 Caring and Sharing Year 2 KSSR Lesson PlanopheliavioletNo ratings yet