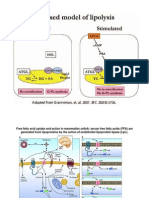
You might also like
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (119)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (265)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (399)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (587)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2219)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5794)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1090)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (344)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (890)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (73)
- Herbal Abortifacient Drugs A ReviewDocument6 pagesHerbal Abortifacient Drugs A Reviewyogesh ushirNo ratings yet
- Infants and ToddlersDocument14 pagesInfants and ToddlersJosias Smith100% (1)
- Psychological Aspects of Stress and Adaptation - AmolDocument12 pagesPsychological Aspects of Stress and Adaptation - Amoldanimon1984No ratings yet
- My Study Plan Guide For AmcDocument7 pagesMy Study Plan Guide For Amc0d&H 8No ratings yet
- Wheel Horse by YearDocument14 pagesWheel Horse by YearNeil SmallwoodNo ratings yet
- Medicinal PlantsDocument211 pagesMedicinal PlantsDr-Abdulrahman GhonemNo ratings yet
- 1.4 Market FailureDocument42 pages1.4 Market FailureRuban PaulNo ratings yet
- Vishnu Sahasra Namam Vol2 PDFDocument301 pagesVishnu Sahasra Namam Vol2 PDFvemuri_sriNo ratings yet
- CMD Civil Shaft Construction Capability Statement 2015Document20 pagesCMD Civil Shaft Construction Capability Statement 2015aecom2009No ratings yet
- Before The Honourable Supreme Court of India: Memorandum On Behalf of PetitonersDocument31 pagesBefore The Honourable Supreme Court of India: Memorandum On Behalf of Petitonerspalkin50% (2)
- Vishnu Sahasra Namam Vol1Document306 pagesVishnu Sahasra Namam Vol1api-3715058100% (2)
- VOLUME VDocument282 pagesVOLUME VMahakaal PanditNo ratings yet
- Anatomy of Lone Wolf Terrorism Special EDocument30 pagesAnatomy of Lone Wolf Terrorism Special EMika RainmanNo ratings yet
- B30 10 Rapid Analysis of Lysine For Feedstuffs ApplicationsDocument3 pagesB30 10 Rapid Analysis of Lysine For Feedstuffs ApplicationsalberiocygnusNo ratings yet
- Vishnusahasranaamam IIT LEctureDocument12 pagesVishnusahasranaamam IIT LEcturealberiocygnusNo ratings yet
- A Freshman Year, Far From Home - Global - The Chronicle of Higher EducationDocument29 pagesA Freshman Year, Far From Home - Global - The Chronicle of Higher EducationalberiocygnusNo ratings yet
- Angiotensin-Converting Enzyme Inhibitors SideeffectsDocument7 pagesAngiotensin-Converting Enzyme Inhibitors SideeffectsalberiocygnusNo ratings yet
- WHO Chapter 6Document8 pagesWHO Chapter 6alberiocygnusNo ratings yet
- Who Factor Foodborne Illness'Document14 pagesWho Factor Foodborne Illness'syooloveNo ratings yet
- WHO Chapter 7Document6 pagesWHO Chapter 7alberiocygnusNo ratings yet
- Asset The MarchDocument6 pagesAsset The MarchalberiocygnusNo ratings yet
- WHO Chapter 7Document6 pagesWHO Chapter 7alberiocygnusNo ratings yet
- Homage to the Nine PlanetsDocument2 pagesHomage to the Nine Planetstempo97217483No ratings yet
- WHO ReferencesDocument4 pagesWHO ReferencesalberiocygnusNo ratings yet
- WHO - Economic Impact of Foodborne IllnessDocument12 pagesWHO - Economic Impact of Foodborne IllnessEm WaNo ratings yet
- ResistDocument101 pagesResistalberiocygnusNo ratings yet
- FA Metabolism Update and HPA EffectsDocument3 pagesFA Metabolism Update and HPA EffectsalberiocygnusNo ratings yet
- ResistDocument101 pagesResistalberiocygnusNo ratings yet
- VWR Drosophila BrochureDocument5 pagesVWR Drosophila BrochurealberiocygnusNo ratings yet
- Homage to the Nine PlanetsDocument2 pagesHomage to the Nine Planetstempo97217483No ratings yet
- Alginate Is Not A Significant Component of The PA EPSDocument6 pagesAlginate Is Not A Significant Component of The PA EPSalberiocygnusNo ratings yet
- C. Elgans-A Practical Appraoch-IAN HOPEDocument125 pagesC. Elgans-A Practical Appraoch-IAN HOPEalberiocygnusNo ratings yet
- Feat 0410 TrendsDocument14 pagesFeat 0410 TrendsalberiocygnusNo ratings yet
- D 2144 - 01 - RdixndqDocument4 pagesD 2144 - 01 - RdixndqjayakumarNo ratings yet
- Changes in Demand and Supply of Face Masks Under CovidDocument3 pagesChanges in Demand and Supply of Face Masks Under CovidHanh HoangNo ratings yet
- Final Final PosterDocument1 pageFinal Final Posterapi-486056653No ratings yet
- Bio Project-Polymerase Chain ReactionDocument21 pagesBio Project-Polymerase Chain ReactionS.AbiniveshNo ratings yet
- Kuratif RacunDocument18 pagesKuratif RacunYsrwncyNo ratings yet
- HPAD 201 Sat Days 2 and 3Document11 pagesHPAD 201 Sat Days 2 and 3Arianne A ZamoraNo ratings yet
- INTUSSUSCEPTIONDocument1 pageINTUSSUSCEPTIONMaecy PasionNo ratings yet
- Caffeine's Effect on Daphnia Heart RateDocument2 pagesCaffeine's Effect on Daphnia Heart RateMianto NamikazeNo ratings yet
- (Tertiary Level Biology) R. J. Wootton (Auth.) - Fish Ecology-Springer Netherlands (1992)Document218 pages(Tertiary Level Biology) R. J. Wootton (Auth.) - Fish Ecology-Springer Netherlands (1992)John Davi JonesNo ratings yet
- The Payment of Bonus Act 1965 PDFDocument30 pagesThe Payment of Bonus Act 1965 PDFappu kunda100% (1)
- Calculation of shell and dish thicknessDocument2 pagesCalculation of shell and dish thicknessThiru AnanthNo ratings yet
- LAOKEN Comparison With J&JDocument3 pagesLAOKEN Comparison With J&JMario Alfonso MartinezNo ratings yet
- The Evolution of FungiDocument15 pagesThe Evolution of Fungi2010eon2010No ratings yet
- Clarifying Questions on the CPRDocument20 pagesClarifying Questions on the CPRmingulNo ratings yet
- MAstering IATFDocument20 pagesMAstering IATFGyanesh_DBNo ratings yet
- XFY1548移动式筛分站术规格书Document16 pagesXFY1548移动式筛分站术规格书abangNo ratings yet
- MANUAL Health O Meter Scale 800KLDocument2 pagesMANUAL Health O Meter Scale 800KLElkin MaldonadoNo ratings yet
- NTTM632 Anand Informatica-ETL Lead ResumeDocument8 pagesNTTM632 Anand Informatica-ETL Lead ResumeMohammed JawadNo ratings yet
- Strain Gauge Load Cells LPB0005IDocument2 pagesStrain Gauge Load Cells LPB0005ILordbyron23No ratings yet
- Wastewater Collection Systems Comparison: William T. Hensley, International Territory Manager, Orenco Systems, IncDocument5 pagesWastewater Collection Systems Comparison: William T. Hensley, International Territory Manager, Orenco Systems, IncmeskbNo ratings yet
- ZCT ZCT ZCT ZCT: 40S 60S 80S 120S 210SDocument1 pageZCT ZCT ZCT ZCT: 40S 60S 80S 120S 210SWilliam TanNo ratings yet