You might also like
- 2012 Japanese Meridian TherapyDocument27 pages2012 Japanese Meridian Therapy1Rogerio100% (7)
- Muscle Injuries: Biology and TreatmentDocument20 pagesMuscle Injuries: Biology and TreatmentZulham Denpasar100% (2)
- A History of Manipulative TherapyDocument10 pagesA History of Manipulative TherapyChiromedBangkokNo ratings yet
- 2012 JOSPT Guidelines LowBackPainDocument58 pages2012 JOSPT Guidelines LowBackPain1RogerioNo ratings yet
- 2012 Myofascial Pain of The Jaw MusclesDocument8 pages2012 Myofascial Pain of The Jaw Muscles1RogerioNo ratings yet
- Appropriate Use of Diagnostic Imaging in LB PainDocument10 pagesAppropriate Use of Diagnostic Imaging in LB Pain1RogerioNo ratings yet
- 2010 Gamma-Band Oscillations in Fronto-Central Areas During Performance of A Sensorio Motor Integration TaskDocument4 pages2010 Gamma-Band Oscillations in Fronto-Central Areas During Performance of A Sensorio Motor Integration Task1RogerioNo ratings yet
- Lesão Condral Da Patela Mar2012-Imaging-LeahyDocument1 pageLesão Condral Da Patela Mar2012-Imaging-Leahy1RogerioNo ratings yet
- The Pearls and Pitfalls of Magnetic Resonance Imaging For The SpineDocument13 pagesThe Pearls and Pitfalls of Magnetic Resonance Imaging For The Spine1RogerioNo ratings yet
- 2012 Strain Hardening of Fascia JBMTDocument7 pages2012 Strain Hardening of Fascia JBMT1RogerioNo ratings yet
- Active Fascial ContractilityDocument5 pagesActive Fascial ContractilitygavinbroomesNo ratings yet
- 2010 The Fascial Manipulation Technique and Its Biomechanical Model: A Guide To The Human Fascial SystemDocument3 pages2010 The Fascial Manipulation Technique and Its Biomechanical Model: A Guide To The Human Fascial System1Rogerio100% (1)
- 2012 Tissue Adaptation To Physical StressDocument23 pages2012 Tissue Adaptation To Physical Stress1RogerioNo ratings yet
- 2003 Fascial Mechanoreceptors and Their Potential Role in Deep Tissue ManipulationDocument10 pages2003 Fascial Mechanoreceptors and Their Potential Role in Deep Tissue Manipulation1RogerioNo ratings yet
- Histological Study of The Deep Fascia of The Upper Limb by Stecco, Porzionato Macchi, Et AlDocument6 pagesHistological Study of The Deep Fascia of The Upper Limb by Stecco, Porzionato Macchi, Et AlTensegrity Wiki100% (1)
- 2008 Vastus Medialis Activation During KneeDocument11 pages2008 Vastus Medialis Activation During Knee1RogerioNo ratings yet
- 2003 Fascial Plasticity - A New Neurobiological Explanation - Part 1 and Part 2Document22 pages2003 Fascial Plasticity - A New Neurobiological Explanation - Part 1 and Part 21Rogerio100% (1)
- Muscle Injuries and Repair: Current Trends in ResearchDocument11 pagesMuscle Injuries and Repair: Current Trends in Research1RogerioNo ratings yet
- 2008 Body As Movement System - A Premise For Structural IntegrationDocument14 pages2008 Body As Movement System - A Premise For Structural Integration1RogerioNo ratings yet
- Muscle Injuries: Biology and TreatmentDocument20 pagesMuscle Injuries: Biology and TreatmentZulham Denpasar100% (2)
- 2006 The European Fascia Research Project by PHD Robert Schleip and TeamDocument8 pages2006 The European Fascia Research Project by PHD Robert Schleip and Team1RogerioNo ratings yet
- 2005 Efeito Anti Infl Do ExerDocument10 pages2005 Efeito Anti Infl Do Exer1RogerioNo ratings yet
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5783)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (890)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (399)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (587)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (265)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (72)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (344)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2219)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (119)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- UN AIDS Miles-To-Go - enDocument268 pagesUN AIDS Miles-To-Go - ensofiabloem100% (1)
- CIPHIDocument256 pagesCIPHIसदानंद देशपांडेNo ratings yet
- The Endocrine Pancreas: Regulation of Carbohydrate MetabolismDocument61 pagesThe Endocrine Pancreas: Regulation of Carbohydrate MetabolismFatima OngNo ratings yet
- The Behavioral Medicine Treatment Planner - Douglas E. DeGood & Angela L. Crawford & Arthur E. JongsmaDocument283 pagesThe Behavioral Medicine Treatment Planner - Douglas E. DeGood & Angela L. Crawford & Arthur E. JongsmaSergio Mora88% (8)
- Ubd Stage 1-3Document24 pagesUbd Stage 1-3api-336167164No ratings yet
- MSA Testing Reveals Body's Energetic HealthDocument4 pagesMSA Testing Reveals Body's Energetic HealthDenise MathreNo ratings yet
- BONE Level 2-BDocument60 pagesBONE Level 2-Bjefri banjarnahorNo ratings yet
- Understanding Mental Illness Across CulturesDocument6 pagesUnderstanding Mental Illness Across Culturesjessaminequeency4128No ratings yet
- How Brain Science Can Inform Architectural DesignDocument12 pagesHow Brain Science Can Inform Architectural DesignadrianmoleavinNo ratings yet
- Nadi Literally MeansDocument9 pagesNadi Literally MeansThomas DürstNo ratings yet
- Spectrophotometric determination of total phenolic content in Gul-e-ZoofaDocument4 pagesSpectrophotometric determination of total phenolic content in Gul-e-Zoofayulyansyah susantoNo ratings yet
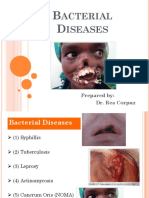
- Acterial Iseases: Prepared By: Dr. Rea CorpuzDocument63 pagesActerial Iseases: Prepared By: Dr. Rea CorpuzMedecine Dentaire100% (2)
- Levorphanol - The Forgotten Opioid PDFDocument6 pagesLevorphanol - The Forgotten Opioid PDFfchem11No ratings yet
- Intraventricular Hemorrhage (IVH) : Intensive Care Nursery House Staff ManualDocument3 pagesIntraventricular Hemorrhage (IVH) : Intensive Care Nursery House Staff Manualjimzz44No ratings yet
- Cranial Base Dan Emergency CaseDocument34 pagesCranial Base Dan Emergency Caserahmad wahyuNo ratings yet
- The Bobath Concept (NDT) As Rehabilitation in Stroke Patients - Revisión SistemáticaDocument8 pagesThe Bobath Concept (NDT) As Rehabilitation in Stroke Patients - Revisión SistemáticaCristian Manuel Goicochea ChuqueNo ratings yet
- Risk Management in Medical LaboratoriesDocument32 pagesRisk Management in Medical Laboratoriesapanisile14142No ratings yet
- Tuberculosis and Typhoid: Deadly Diseases in the PhilippinesDocument37 pagesTuberculosis and Typhoid: Deadly Diseases in the PhilippinesAriel FuentespinaNo ratings yet
- Male Stress Urinary Incontinence: Giulio Del Popolo Donatella Pistolesi Vincenzo Li MarziDocument189 pagesMale Stress Urinary Incontinence: Giulio Del Popolo Donatella Pistolesi Vincenzo Li MarziVeronika AtaNo ratings yet
- Datex-Ohmeda Aespire 7900 - Maintenance and Troubleshooting PDFDocument118 pagesDatex-Ohmeda Aespire 7900 - Maintenance and Troubleshooting PDFJosé Morales Echenique100% (1)
- Oral Physio FinalDocument34 pagesOral Physio FinaldraggarwalNo ratings yet
- Miocarditis Por Clozapina Ronaldson2017 Revisión SistematicaDocument19 pagesMiocarditis Por Clozapina Ronaldson2017 Revisión SistematicaAlejandro PiscitelliNo ratings yet
- Pediatric Surgical Handbook PDFDocument139 pagesPediatric Surgical Handbook PDFNia LahidaNo ratings yet
- And Technical Description: User ManualDocument100 pagesAnd Technical Description: User ManualYouness Ben TibariNo ratings yet
- Drugs & Cosmetics ActDocument70 pagesDrugs & Cosmetics ActAnonymous ibmeej9No ratings yet
- Turmeric Benefits: Health Benefits of Turmeric - Are There Any Side Effects of Turmeric?Document5 pagesTurmeric Benefits: Health Benefits of Turmeric - Are There Any Side Effects of Turmeric?Angelyn Taberna NatividadNo ratings yet
- Bench MarkingDocument16 pagesBench MarkingNorj BaraniNo ratings yet
- Pearls in The Management of ExotropiaDocument4 pagesPearls in The Management of ExotropiaGuy BarnettNo ratings yet
- Nutrition Quiz: Build a Healthy DietDocument17 pagesNutrition Quiz: Build a Healthy DietJan Angelo OcadoNo ratings yet
- Wound Management ProcedureDocument51 pagesWound Management ProcedureSisca Dwi AgustinaNo ratings yet