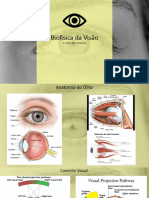
You might also like
- VisãoDocument50 pagesVisãoNathalia AzuosNo ratings yet
- Morfofisiologia Da VisaoDocument55 pagesMorfofisiologia Da VisaoReinaldo RiquetNo ratings yet
- BiofisicaDocument17 pagesBiofisicarmdsNo ratings yet
- Órgãos Dos Sentidos - VisãoDocument9 pagesÓrgãos Dos Sentidos - VisãoClara Ventura100% (2)
- 8 Olho e Visao HumanaDocument22 pages8 Olho e Visao HumanaAndré Martins CésarNo ratings yet
- Sequenciamento Completo Do Genoma: Diferenciando entre organismos, precisamente, como nunca antesFrom EverandSequenciamento Completo Do Genoma: Diferenciando entre organismos, precisamente, como nunca antesRating: 5 out of 5 stars5/5 (1)
- Programa Visão RestauradaDocument17 pagesPrograma Visão RestauradaSuporteNo ratings yet
- Aula 3 Fisiologia Do Sistema NervosoDocument24 pagesAula 3 Fisiologia Do Sistema NervosoEdiane AlbuquerqueNo ratings yet
- Sistema MuscularDocument43 pagesSistema MuscularIvanCostaPassosNo ratings yet
- Exame Físico Ombro, Cotovelo, Mão SlideDocument70 pagesExame Físico Ombro, Cotovelo, Mão SlideDayanna DheniferNo ratings yet
- Exercícios AcomodativosDocument30 pagesExercícios AcomodativosShalom Adonai0% (1)
- Apostila Órgãos Dos SentidosDocument6 pagesApostila Órgãos Dos SentidosMauricio LimaNo ratings yet
- Experiências e Trajetórias da Massoterapia no Instituto Federal do Paraná: Campus LondrinaFrom EverandExperiências e Trajetórias da Massoterapia no Instituto Federal do Paraná: Campus LondrinaNo ratings yet
- Retinopatia DiabéticaDocument16 pagesRetinopatia DiabéticaRUTHNo ratings yet
- Bem viver: 10 maneiras práticas de tornar sua vida melhor e mais saudávelFrom EverandBem viver: 10 maneiras práticas de tornar sua vida melhor e mais saudávelNo ratings yet
- O Olho HumanoDocument17 pagesO Olho Humanoalexanr2232No ratings yet
- MiologiaDocument19 pagesMiologiaJeniffer Rosa MartinsNo ratings yet
- Aula 01Document13 pagesAula 01Larissa SousaNo ratings yet
- Apostila 03 1Document33 pagesApostila 03 1pecceriniNo ratings yet
- Artigo 060706Document3 pagesArtigo 060706Nilo José BNo ratings yet
- EpicondiliteDocument16 pagesEpicondiliteNatsume LiraNo ratings yet
- Anatomia 2Document26 pagesAnatomia 2Belmiro G. White LwambaNo ratings yet
- 16 - AntiparkinsonianosDocument6 pages16 - AntiparkinsonianosForwomenNo ratings yet
- Esparmo MuscularDocument10 pagesEsparmo MuscularMaíra PiresNo ratings yet
- 71 - A ImportYncia Das Manobras Gerais Da Quiropraxia em Pacientes Com HYrnia de DiscoDocument15 pages71 - A ImportYncia Das Manobras Gerais Da Quiropraxia em Pacientes Com HYrnia de DiscoCesar Odilon PereiraNo ratings yet
- Órgãos Dos SentidosDocument32 pagesÓrgãos Dos SentidosRodrigo de Oliveira ReisNo ratings yet
- Raio XDocument26 pagesRaio XAllan ThopsonNo ratings yet
- Aula 4 - Olho HumanoDocument68 pagesAula 4 - Olho HumanoJoão LoteNo ratings yet
- Estudo Dirigido - VisãoDocument4 pagesEstudo Dirigido - Visãocarlos gabrielNo ratings yet
- Material Saúde OcularDocument22 pagesMaterial Saúde OcularJoãoRicardoPanzera100% (1)
- E Book FIBROSE PÓS CIRURGIA PLASTICADocument19 pagesE Book FIBROSE PÓS CIRURGIA PLASTICAPaty SilvaNo ratings yet
- Aula 3 - Terapia Manual PompagemDocument39 pagesAula 3 - Terapia Manual PompagemALAMY ANISIO DOS SANTOSNo ratings yet
- Cosmetologia - TecnologiaDocument93 pagesCosmetologia - TecnologiamanuelNo ratings yet
- Biomex MusculosDocument150 pagesBiomex MusculosFernanda GonçalvesNo ratings yet
- Apostila Fisio Patologia Do Lin Fedem ADocument7 pagesApostila Fisio Patologia Do Lin Fedem AGladnejpNo ratings yet
- Bursite Do Olécrano Ou Bursite Do CotoveloDocument5 pagesBursite Do Olécrano Ou Bursite Do CotoveloPaulo Monteiro100% (1)
- Aula 02 - Lesões Nervosas PeriféricasDocument67 pagesAula 02 - Lesões Nervosas PeriféricasMaya Silveira100% (1)
- Sono NREM e Sono REM PDFDocument3 pagesSono NREM e Sono REM PDFJehonnesNo ratings yet
- Degeneração Macular, Tratamento Com PinholeDocument5 pagesDegeneração Macular, Tratamento Com PinholeMarco GuimaraesNo ratings yet
- Membro FantasmaDocument23 pagesMembro FantasmaAngelita CarvalhoNo ratings yet
- 6 Sistema Nervoso SensorialDocument40 pages6 Sistema Nervoso SensorialWander Junior100% (1)
- Estudo Dirigido de Fisiologia 1 - Com RespostasDocument4 pagesEstudo Dirigido de Fisiologia 1 - Com RespostasThaisNo ratings yet
- Patologia Dos Anexos OcularesDocument16 pagesPatologia Dos Anexos OculareswendellgiordaniNo ratings yet
- Cintilografia ÓsseaDocument8 pagesCintilografia ÓsseaPedro Henrique DoniniNo ratings yet
- Avaliação Dos Nervos CranianosDocument1 pageAvaliação Dos Nervos CranianosAdriele TaianeNo ratings yet
- Nova Roda Dos AlimentosDocument1 pageNova Roda Dos AlimentosPaula CorreiaNo ratings yet
- Guia AlimentarDocument35 pagesGuia Alimentarjpedro88100% (1)
- Paul Feyerabend - Trabalho FinalDocument2 pagesPaul Feyerabend - Trabalho FinalPaula CorreiaNo ratings yet
- Histologia - Poster FINALDocument1 pageHistologia - Poster FINALPaula CorreiaNo ratings yet
- Envelhecimento Celucar e A Importancia Da Vitamina CDocument16 pagesEnvelhecimento Celucar e A Importancia Da Vitamina CPaula CorreiaNo ratings yet
- Plano de Aula para Atividade Discursiva de Educação Alimentar - Com Figuras-Pronto-RevisadoDocument8 pagesPlano de Aula para Atividade Discursiva de Educação Alimentar - Com Figuras-Pronto-RevisadoCHEILA50% (2)
- Cânon BíblicoDocument12 pagesCânon Bíblicorodrigoepl7411No ratings yet
- Edgar Allan Poe - A Queda Da Casa de UsherDocument13 pagesEdgar Allan Poe - A Queda Da Casa de UsherpehdefigoNo ratings yet
- A Grande Aventura de Maria Fumaça PDFDocument1 pageA Grande Aventura de Maria Fumaça PDFTatiane GonçalvesNo ratings yet
- Lista de Inscritos A Curso: Democracia DigitalDocument14 pagesLista de Inscritos A Curso: Democracia DigitalPartido Liberal ColombianoNo ratings yet
- Bonetti LucasZangirolami DDocument563 pagesBonetti LucasZangirolami DGustavo KraemerNo ratings yet
- TESE DOUTORADO LUIS LEITE Versao Internet PDFDocument152 pagesTESE DOUTORADO LUIS LEITE Versao Internet PDFArtur OrtenbladNo ratings yet
- Anais Terceiro Colóquio Internacional FITADocument103 pagesAnais Terceiro Colóquio Internacional FITAIgor GomesNo ratings yet
- Origem Do HalloweenDocument2 pagesOrigem Do HalloweenShirley DaianeNo ratings yet
- Bernard Tschumi Seis Conceitos Arte e Ensaios UFRJ PDFDocument14 pagesBernard Tschumi Seis Conceitos Arte e Ensaios UFRJ PDFWilton MedeirosNo ratings yet
- Apocalipse - Estudo 06 - A Visão Do Livro e Dos SelosDocument3 pagesApocalipse - Estudo 06 - A Visão Do Livro e Dos SelosgguerinoNo ratings yet
- Rolamentos Autocompensadores de Rolos: DimensõesDocument4 pagesRolamentos Autocompensadores de Rolos: DimensõesdoidobskNo ratings yet
- Prova Especial.2.6anoDocument4 pagesProva Especial.2.6anoJerônimo MedeirosNo ratings yet
- Orla DenteadaDocument3 pagesOrla DenteadaFernando Pozza33% (3)
- Teste 5ºDocument5 pagesTeste 5ºFamília Feliz100% (1)
- Revista Capriccho - Simpo2020 PDFDocument6 pagesRevista Capriccho - Simpo2020 PDFMarcos AraújoNo ratings yet
- Caracterização Cultural de AveiroDocument5 pagesCaracterização Cultural de Aveiroalexandra SilvaNo ratings yet
- Nicolau MaquiavelDocument6 pagesNicolau MaquiavelJhonny PetrickNo ratings yet
- Avaliação Dom Casmurro 01Document7 pagesAvaliação Dom Casmurro 01Ado MachadoNo ratings yet
- Cifra Club - ELA É CARIOCA - Adriana Calcanhotto (Cifra para Violão e Guitarra)Document3 pagesCifra Club - ELA É CARIOCA - Adriana Calcanhotto (Cifra para Violão e Guitarra)Isådora de CåstroNo ratings yet
- Frithjof Schuon A Base Da Religiao e Da MetafisicaDocument10 pagesFrithjof Schuon A Base Da Religiao e Da MetafisicaRose Carmo100% (1)
- Xamanismo - Animais de PoderDocument4 pagesXamanismo - Animais de PoderminomarNo ratings yet
- Oexp12 Ficha Ed Lit Ano Morte Ricardo ReisDocument1 pageOexp12 Ficha Ed Lit Ano Morte Ricardo ReisSofia Valerio100% (1)
- Bronzeamento Artificial Com o PhotoshopDocument2 pagesBronzeamento Artificial Com o PhotoshopAlexandre NascimentoNo ratings yet
- A Arte de Argumentar Gerenciando Razao e EmocaoDocument7 pagesA Arte de Argumentar Gerenciando Razao e EmocaoAna Paula BarrosNo ratings yet
- O Livro Como Objeto Da ArteDocument53 pagesO Livro Como Objeto Da ArteFelipe TognoliNo ratings yet
- Fichamento - As Palavras e As CoisasDocument2 pagesFichamento - As Palavras e As CoisaslchaussardNo ratings yet
- Material para Download !!!!!!!Document7 pagesMaterial para Download !!!!!!!gato0% (1)
- 31 G 2Document35 pages31 G 2cleitoneqNo ratings yet
- Batalha Geométrica RecicladaDocument4 pagesBatalha Geométrica RecicladaRobson Santana SNo ratings yet