You might also like
- Guia de Vigilancia EpidemiologicaDocument523 pagesGuia de Vigilancia EpidemiologicaNílvia Daniele Ferreira100% (2)
- ADS no domicílio: relato de experiência psicológicaDocument14 pagesADS no domicílio: relato de experiência psicológicaAndyara OliveiraNo ratings yet
- Introduzindo a perspectiva de gênero na formação docenteDocument15 pagesIntroduzindo a perspectiva de gênero na formação docenteJoão Paulo AndradeNo ratings yet
- A Gestao Ambiental Na Instalacao de Plataformas OffshoreDocument41 pagesA Gestao Ambiental Na Instalacao de Plataformas OffshoreRafael CassimiroNo ratings yet
- Educação Infantil e As Práticas de Cuidar e Educar No Contexto DasDocument20 pagesEducação Infantil e As Práticas de Cuidar e Educar No Contexto DasLuciano DinizNo ratings yet
- Conceitos Fundamentais PDFDocument2 pagesConceitos Fundamentais PDFPaulo VictorNo ratings yet
- Fotografia Meio e Linguagem Dentro Da ModaDocument76 pagesFotografia Meio e Linguagem Dentro Da ModaJ Paulo CoutinhoNo ratings yet
- Filosofia saber críticoDocument70 pagesFilosofia saber críticoMANOEL MESSIAS ARAUJONo ratings yet
- A importância das artes visuais no ensinoDocument10 pagesA importância das artes visuais no ensinoStela Maris Britto MazieroNo ratings yet
- Tiristores e Retificadores Control A DosDocument20 pagesTiristores e Retificadores Control A DosBuzioliNo ratings yet
- Tipos e Aplicações de CaldeiraDocument10 pagesTipos e Aplicações de CaldeiraJosi AlvarengaNo ratings yet
- O Prazer de Aprender Com A Informática Na Educação InfantilDocument16 pagesO Prazer de Aprender Com A Informática Na Educação InfantilEddie HermanoNo ratings yet
- Ações de Prevenção Primária e Secundária No Controle Do CâncerDocument66 pagesAções de Prevenção Primária e Secundária No Controle Do CâncerQualis ConsultoriaNo ratings yet
- Código de Ética Do Guia de TurismoDocument2 pagesCódigo de Ética Do Guia de TurismoMaríliaGonçalvesNo ratings yet
- Normas Técnicas para Produção de Texto em BrailleDocument73 pagesNormas Técnicas para Produção de Texto em BrailleAtaniel CampeloNo ratings yet
- Qsms Qualidade Saude Meio Ambiente SegurancaDocument117 pagesQsms Qualidade Saude Meio Ambiente SegurancaOthon MonteiroNo ratings yet
- Como Funciona A Geração de Imagens Por Ressonância MagnéticaDocument10 pagesComo Funciona A Geração de Imagens Por Ressonância MagnéticaDenise Leite da SilvaNo ratings yet
- PRO - DISTRIBU-ENGE-0019 - Manual de Procedimentos Operacionais Padrão - Rede de Distribuição Até 34,5 KV - Utilizando Linha Viva PDFDocument65 pagesPRO - DISTRIBU-ENGE-0019 - Manual de Procedimentos Operacionais Padrão - Rede de Distribuição Até 34,5 KV - Utilizando Linha Viva PDFAlex MeirelesNo ratings yet
- Layout de Sites e Botões AnimadosDocument6 pagesLayout de Sites e Botões AnimadosTakuma SukeNo ratings yet
- AjustagemDocument18 pagesAjustagemVictor HugoNo ratings yet
- Prospecção de Cenário No Varejo Farmacêutico BrasileiroDocument24 pagesProspecção de Cenário No Varejo Farmacêutico BrasileiroMarcelo Dos SantosNo ratings yet
- 1 - Correspondência Entre Comandos Do WORD 2003 e 2007Document75 pages1 - Correspondência Entre Comandos Do WORD 2003 e 2007Manoel HigorNo ratings yet
- GerenciamentoRecepçãoHotéisDocument30 pagesGerenciamentoRecepçãoHotéisRenata CristinaNo ratings yet
- AnemiaDocument29 pagesAnemiaPrici SaNo ratings yet
- Manual Do CFOPDocument11 pagesManual Do CFOPMaurício CunhaNo ratings yet
- v2 Celulite Artigo de Revisao PDFDocument6 pagesv2 Celulite Artigo de Revisao PDFJanneNo ratings yet
- Desafios Do Sistema de Segurança Pública No BrasilDocument70 pagesDesafios Do Sistema de Segurança Pública No BrasilIandra MarquesNo ratings yet
- Morfologia e BotânicaDocument26 pagesMorfologia e BotânicaPedro Resende FilhoNo ratings yet
- Processo produtivo farmacêuticoDocument31 pagesProcesso produtivo farmacêuticoRafael DiasNo ratings yet
- Introducao AviculturaDocument92 pagesIntroducao AviculturaGuilherme AndradeNo ratings yet
- Classificações de Políticas de Segurança PúblicaDocument13 pagesClassificações de Políticas de Segurança Públicatilovec890No ratings yet
- A Eficácia Da Lei de Responsabilidade Fiscal Na Gestão Dos Recursos Públicos MunicipaisDocument16 pagesA Eficácia Da Lei de Responsabilidade Fiscal Na Gestão Dos Recursos Públicos Municipaistensol20037159No ratings yet
- Gestão de Facilidades: Robson Quinello José Roberto NicolettiDocument17 pagesGestão de Facilidades: Robson Quinello José Roberto Nicolettileomariano21No ratings yet
- Balanceamento de Ventiladores IndustriaisDocument9 pagesBalanceamento de Ventiladores IndustriaisualassefernandoNo ratings yet
- Caso Usaflex PDFDocument29 pagesCaso Usaflex PDFMISTY EYEDNo ratings yet
- Introdução à Recuperação Sem PinturaDocument28 pagesIntrodução à Recuperação Sem PinturaClaudio FrancelinoNo ratings yet
- 1 - Cristologia (Introdução)Document14 pages1 - Cristologia (Introdução)Carlos PinheiroNo ratings yet
- Conceitos Básicos Sobre Avaliação Das AprendizagensDocument17 pagesConceitos Básicos Sobre Avaliação Das Aprendizagenslulinhaalmeida100% (1)
- Construção Do Círculo CromáticoDocument13 pagesConstrução Do Círculo CromáticoJosé Nunes LadeiraNo ratings yet
- Parasitologia Clinica IntroduçãoDocument46 pagesParasitologia Clinica IntroduçãoSabrina MendesNo ratings yet
- Ensino da Gramática e LinguagemDocument12 pagesEnsino da Gramática e LinguagemAlan Arp100% (1)
- Curso de Contabilidade Geral para Concursos PúblicosDocument183 pagesCurso de Contabilidade Geral para Concursos PúblicosCaroline Varela100% (4)
- O mercado da ovinocultura no BrasilDocument21 pagesO mercado da ovinocultura no BrasilJeferson Silva CarvalhoNo ratings yet
- Cana de Acucar 2020Document66 pagesCana de Acucar 2020Nyelly CoutrimNo ratings yet
- O Tecido Adiposo como Regulador MetabólicoDocument14 pagesO Tecido Adiposo como Regulador MetabólicoKeeyla S.No ratings yet
- OTecido Adiposo Como Centro Reguladordo MetabolismoDocument14 pagesOTecido Adiposo Como Centro Reguladordo Metabolismoalexsandra santosNo ratings yet
- 5e3f9f2f-c181-462e-b2f9-3cd78a6329b9Document69 pages5e3f9f2f-c181-462e-b2f9-3cd78a6329b9bfontesnutriNo ratings yet
- O Tecido Adiposo Como Centro Regulador Do MetabolismoDocument13 pagesO Tecido Adiposo Como Centro Regulador Do Metabolismobumbim0No ratings yet
- 3033 Outro 6267 1 10 20191118Document5 pages3033 Outro 6267 1 10 20191118amanda.macugliaNo ratings yet
- UntitledDocument17 pagesUntitledAline OliveiraNo ratings yet
- Tecidos adiposos: cores e versatilidadeDocument12 pagesTecidos adiposos: cores e versatilidadeYara ZaninNo ratings yet
- O que é metabolismo e como funciona o catabolismo da glicoseDocument13 pagesO que é metabolismo e como funciona o catabolismo da glicoseBeatriz TiveronNo ratings yet
- Dislipidemias: conceito, epidemiologia e metabolismo lipídicoDocument28 pagesDislipidemias: conceito, epidemiologia e metabolismo lipídicoRoberta Violante100% (1)
- Integração MetabólicaDocument8 pagesIntegração MetabólicaAmanda YuriNo ratings yet
- Simulado BIOQUÍMICADocument14 pagesSimulado BIOQUÍMICAMatheusNo ratings yet
- Trabalho N1 Bioquímica - CORRIGIDODocument11 pagesTrabalho N1 Bioquímica - CORRIGIDOJulia MoraesNo ratings yet
- Tecido adiposo: função endócrina e termogênicaDocument32 pagesTecido adiposo: função endócrina e termogênicaGustavo MachadoNo ratings yet
- As adipocinas: hormônios secretados pelo tecido adiposoDocument3 pagesAs adipocinas: hormônios secretados pelo tecido adiposoRobertoNo ratings yet
- Texto 11Document19 pagesTexto 11Milena TessmannNo ratings yet
- Prolactina e Diabetes Melito do tipo 2: o efeito protetor de um hormônio sobre o metabolismo glicídicoFrom EverandProlactina e Diabetes Melito do tipo 2: o efeito protetor de um hormônio sobre o metabolismo glicídicoNo ratings yet
- Livrocrbm 040509Document76 pagesLivrocrbm 040509marisademoura100% (1)
- UFERSA Laboratório QuímicaDocument2 pagesUFERSA Laboratório QuímicaCecília RodriguesNo ratings yet
- Edital Do Concurso Do INSSDocument30 pagesEdital Do Concurso Do INSSBlogdosConcursosNo ratings yet
- Biossegurança em Laboratórios Biomédicos e de Microbiologia PDFDocument290 pagesBiossegurança em Laboratórios Biomédicos e de Microbiologia PDFMylleSilvaNo ratings yet
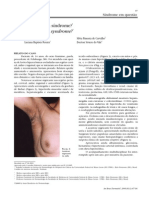
- Síndrome de Berardinelli-SeipDocument4 pagesSíndrome de Berardinelli-SeipguidabiomedicinaNo ratings yet
- Instr010203 v03Document32 pagesInstr010203 v03guidabiomedicinaNo ratings yet
- MB007QDocument13 pagesMB007QguidabiomedicinaNo ratings yet
- Lei do Estatuto do Desarmamento comentadaDocument28 pagesLei do Estatuto do Desarmamento comentadaLeonardo BarreirasNo ratings yet
- Apostila de FisicaDocument59 pagesApostila de FisicaCristiana MarinhoNo ratings yet
- Sistema Nervoso AutônomoDocument20 pagesSistema Nervoso AutônomoTarcilio Araujo Brito100% (1)
- Avaliação Por Imagem Da Área de Gordura Visceral e SuasDocument7 pagesAvaliação Por Imagem Da Área de Gordura Visceral e SuasguidabiomedicinaNo ratings yet
- Adipocitocinas e suas ações no metabolismo e sistema vascularDocument12 pagesAdipocitocinas e suas ações no metabolismo e sistema vascularguidabiomedicina100% (1)
- Ensinamentos de Central do Brasil sobre educação e políticas públicasDocument16 pagesEnsinamentos de Central do Brasil sobre educação e políticas públicasThamires LeãoNo ratings yet
- Membros InferioresDocument26 pagesMembros InferioresPhelipe AquinoNo ratings yet
- A Participação Do Tecido Adiposo Visceral Na Gênese Da Hipertensão e Doença Cardiovascular Aterogênica. Um Conceito EmergenteDocument13 pagesA Participação Do Tecido Adiposo Visceral Na Gênese Da Hipertensão e Doença Cardiovascular Aterogênica. Um Conceito Emergentejhot7No ratings yet
- Montagem de cariograma humanoDocument5 pagesMontagem de cariograma humanoguidabiomedicina100% (1)
- Diagnostico Integrado em OrtopediaDocument26 pagesDiagnostico Integrado em OrtopediaBira ViannaNo ratings yet
- ARQUIVO Artigo SNHCDocument11 pagesARQUIVO Artigo SNHCguidabiomedicinaNo ratings yet
- 03 TecnicascitogeneticasDocument17 pages03 TecnicascitogeneticasVinicius DionisioNo ratings yet
- RetificacaoeditaldepartbahiapoliciaDocument9 pagesRetificacaoeditaldepartbahiapoliciaguidabiomedicinaNo ratings yet
- Ag AcDocument32 pagesAg AcSoninha MotaNo ratings yet
- Afroasia n16 p54Document16 pagesAfroasia n16 p54guidabiomedicinaNo ratings yet
- Roteiro Projeto Pesquisa RelatorioDocument33 pagesRoteiro Projeto Pesquisa Relatoriorafagz13No ratings yet
- Biologia: Glossário de termosDocument92 pagesBiologia: Glossário de termosguidabiomedicina0% (1)
- 2006dez Forense1Document11 pages2006dez Forense1Wiliam SabinoNo ratings yet
- Biologia: Glossário de termosDocument92 pagesBiologia: Glossário de termosguidabiomedicina0% (1)
- Livro Zootecnia Com Sustentabilidade - Formato E-BookDocument121 pagesLivro Zootecnia Com Sustentabilidade - Formato E-BooklrribeiroNo ratings yet
- Hemocomponentes e hemoderivados: indicações e aplicaçõesDocument5 pagesHemocomponentes e hemoderivados: indicações e aplicaçõesMichelle OliveiraNo ratings yet
- Deriva GenéticaDocument5 pagesDeriva GenéticaKlaudio MagalhãesNo ratings yet
- Exercícios para fortalecer a Glândula TIMODocument13 pagesExercícios para fortalecer a Glândula TIMOJoara GuimarãesNo ratings yet
- 47 - Os BenefYcios Da AplicaYYo Da Luz Intensa Pulsada Na EpilaYYo e em Diversas AfecYes Na Pele - RLDocument13 pages47 - Os BenefYcios Da AplicaYYo Da Luz Intensa Pulsada Na EpilaYYo e em Diversas AfecYes Na Pele - RLwwsomNo ratings yet
- SifilisDocument6 pagesSifilisFernanda FurlanNo ratings yet
- ManuaNutrologia AlimentacaoDocument152 pagesManuaNutrologia AlimentacaoMagali Souza Chef VilleNo ratings yet
- 10 Remédios Que Têm Efeitos Colaterais AssustadoresDocument2 pages10 Remédios Que Têm Efeitos Colaterais Assustadoresx_menxavier3236No ratings yet
- Como Se Preparar Psicologicamente para Provas de Vestibulares e ConcursosDocument4 pagesComo Se Preparar Psicologicamente para Provas de Vestibulares e ConcursosEmerson EliasNo ratings yet
- Grade Curricular FarmáciaDocument16 pagesGrade Curricular FarmáciaFernando MunizNo ratings yet
- Licenciatura Enfermagem ESENFC Bioquímica ClonagemDocument85 pagesLicenciatura Enfermagem ESENFC Bioquímica ClonagemHelena LázaroNo ratings yet
- VitaminasDocument2 pagesVitaminasAdão Marcos GracianoNo ratings yet
- 3-Saúde No Baralho CiganoDocument6 pages3-Saúde No Baralho CiganoDelcilene100% (1)
- LannateBR BulaDocument10 pagesLannateBR BulageovaneNo ratings yet
- Cultivo de plantas medicinaisDocument34 pagesCultivo de plantas medicinaisLarissa Barboza100% (2)
- Jogo Sipat 2Document8 pagesJogo Sipat 2Isabella AmaralNo ratings yet
- 5 Maneiras Que o Alcool Atrapalha A HipertrofiaDocument4 pages5 Maneiras Que o Alcool Atrapalha A HipertrofiaRafael NetoNo ratings yet
- Artrópodes de interesse médico: moscas e miíasesDocument4 pagesArtrópodes de interesse médico: moscas e miíasesLeticia FariasNo ratings yet
- ILSI Livro Carboidratos Glicemia e Saude ONLINEDocument49 pagesILSI Livro Carboidratos Glicemia e Saude ONLINEerikaNo ratings yet
- Conteúdo Da Prova para Monitoria de MicologiaDocument10 pagesConteúdo Da Prova para Monitoria de MicologiaHelena MedradoNo ratings yet
- Doença da Vaca LoucaDocument18 pagesDoença da Vaca LoucaGabriela Almeida100% (1)
- O Legado de Florence NightingaleDocument3 pagesO Legado de Florence NightingaleRaphael Fuga100% (1)
- TCC Final Ana CatarinaDocument29 pagesTCC Final Ana Catarinaandynho16araujoNo ratings yet
- Cervicite FolicularDocument1 pageCervicite FolicularKetlyn OliveiraNo ratings yet
- DNA e Síntese ProteicaDocument5 pagesDNA e Síntese ProteicamanuelafonsecaNo ratings yet
- Aula de Introdução A Fisiologia GeralDocument19 pagesAula de Introdução A Fisiologia GeralMichelle MonalizaNo ratings yet
- Os Cavernomas Cerebrais: Sintomas, Diagnóstico e TratamentoDocument31 pagesOs Cavernomas Cerebrais: Sintomas, Diagnóstico e TratamentoRonaldoSilvaJrNo ratings yet
- A Liberação Miofascial É Uma Técnica Do Método Americano RolfingDocument2 pagesA Liberação Miofascial É Uma Técnica Do Método Americano RolfingMarcelo JotaNo ratings yet
- Aula 5 - Heranças Monogênicas e PoligênicasDocument25 pagesAula 5 - Heranças Monogênicas e PoligênicasCleonete Ferreira De Araujo100% (2)
- Dor lombar e acupunturaDocument16 pagesDor lombar e acupunturaRonisson Guimaraes100% (2)