You might also like
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- 267alphastatus EnglishDocument15 pages267alphastatus EnglishRamon SoaresNo ratings yet
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (587)
- Ulves Jagt På Elg Og Rådyr I Sverige Og Norge Og Byttedyrenes Forsvarsadfærd. Videnskabelig Artikel 2009.Document12 pagesUlves Jagt På Elg Og Rådyr I Sverige Og Norge Og Byttedyrenes Forsvarsadfærd. Videnskabelig Artikel 2009.Ulv i DanmarkNo ratings yet
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (890)
- Shoot, Shovel and Shut Up: Poaching Slows Restoration of Wolf in Scandinavia (Videnskabelig Artikel 2011)Document7 pagesShoot, Shovel and Shut Up: Poaching Slows Restoration of Wolf in Scandinavia (Videnskabelig Artikel 2011)Ulv i DanmarkNo ratings yet
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- Undersøgelse Fra Hviderusland Af, Hvad Der Sker Med Ulvens Diæt, Når de Vilde Byttedyr Forsvinder For Derefter at Komme Igen (Videnskabelig Artikel)Document9 pagesUndersøgelse Fra Hviderusland Af, Hvad Der Sker Med Ulvens Diæt, Når de Vilde Byttedyr Forsvinder For Derefter at Komme Igen (Videnskabelig Artikel)Ulv i DanmarkNo ratings yet
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (399)
- Ulves Sommerjagt På Elge Og Andre Byttedyr I Sverige Og Norge. Videnskabelig Artikel 2008.Document12 pagesUlves Sommerjagt På Elge Og Andre Byttedyr I Sverige Og Norge. Videnskabelig Artikel 2008.Ulv i DanmarkNo ratings yet
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (73)
- Wildlife Comeback in Europe: The Recovery of Selected Mammal and Bird Species. Videnskabelig Rapport 2013.Document312 pagesWildlife Comeback in Europe: The Recovery of Selected Mammal and Bird Species. Videnskabelig Rapport 2013.Ulv i Danmark100% (1)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5794)
- Genintroduktionen Af Ulv Til Yellowstone Nationalpark (Videnskabelig Artikel)Document13 pagesGenintroduktionen Af Ulv Til Yellowstone Nationalpark (Videnskabelig Artikel)Ulv i DanmarkNo ratings yet
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- Norsk Hunulv Vandrer Over 10.000 KM På To År (Videnskabelig Artikel 2007)Document5 pagesNorsk Hunulv Vandrer Over 10.000 KM På To År (Videnskabelig Artikel 2007)Ulv i DanmarkNo ratings yet
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- Prey Choice and Diet of Wolves Related To Ungulate Communities and Wolf Subpopulations in Poland (Videnskabelig Artikel 2012)Document14 pagesPrey Choice and Diet of Wolves Related To Ungulate Communities and Wolf Subpopulations in Poland (Videnskabelig Artikel 2012)Ulv i DanmarkNo ratings yet
- Wolf Howling Is Mediated by Relationship Quality Rather Than Underlying Emotional Stress (Videnskabelig Artikel 2013)Document4 pagesWolf Howling Is Mediated by Relationship Quality Rather Than Underlying Emotional Stress (Videnskabelig Artikel 2013)Ulv i DanmarkNo ratings yet
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- Grey Wolf Subsidies To Scavengers in Yellowstone National Park (Videnskabelig Artikel)Document9 pagesGrey Wolf Subsidies To Scavengers in Yellowstone National Park (Videnskabelig Artikel)Ulv i DanmarkNo ratings yet
- Undersøgelse Af Ulve Med Skab I Rocky Mountains, USA (Videnskabelig Artikel 2010)Document6 pagesUndersøgelse Af Ulve Med Skab I Rocky Mountains, USA (Videnskabelig Artikel 2010)Ulv i DanmarkNo ratings yet
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- Ulv Long Distance Dispersal of Rescued Wolf From N Apennines To W Alps Ciucci Et Al 2009Document8 pagesUlv Long Distance Dispersal of Rescued Wolf From N Apennines To W Alps Ciucci Et Al 2009Ulv i DanmarkNo ratings yet
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1090)
- DNA-analyse Af de Polske Og Tyske Ulve (Videnskabelig Artikel 2013)Document16 pagesDNA-analyse Af de Polske Og Tyske Ulve (Videnskabelig Artikel 2013)Ulv i DanmarkNo ratings yet
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2219)
- Ulvenes Hyleadfærd I Polen (Videnskabelig Artikel 2007)Document7 pagesUlvenes Hyleadfærd I Polen (Videnskabelig Artikel 2007)Ulv i DanmarkNo ratings yet
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- LIFE and Human Coexistence With Large Carnivores, EU 2013 (Rapport)Document76 pagesLIFE and Human Coexistence With Large Carnivores, EU 2013 (Rapport)Ulv i DanmarkNo ratings yet
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (344)
- Status, Management and Distribution of Large Carnivores - Bear, Lynx, Wolf & Wolverine - in Europe (Marts 2013)Document72 pagesStatus, Management and Distribution of Large Carnivores - Bear, Lynx, Wolf & Wolverine - in Europe (Marts 2013)Ulv i DanmarkNo ratings yet
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (265)
- Undersøgelse Af Ulve Med Skab I Yellowstone (Videnskabelig Artikel 2012)Document13 pagesUndersøgelse Af Ulve Med Skab I Yellowstone (Videnskabelig Artikel 2012)Ulv i DanmarkNo ratings yet
- Undersøgelse Fra Spanien Af Skab Hos Ulv, Ræv, Rådyr, Kronhjort Og Gemse (Videnskabelig Artikel 2012)Document4 pagesUndersøgelse Fra Spanien Af Skab Hos Ulv, Ræv, Rådyr, Kronhjort Og Gemse (Videnskabelig Artikel 2012)Ulv i DanmarkNo ratings yet
- IUCN Wolf Specialist Group Manifesto On Wolf Conservation (2000)Document4 pagesIUCN Wolf Specialist Group Manifesto On Wolf Conservation (2000)Ulv i DanmarkNo ratings yet
- Undersøgelse Af Ulves Påvirkning Af Hjortes Fitness Og Drægtighed. Videnskabelig Artikel 2013.Document8 pagesUndersøgelse Af Ulves Påvirkning Af Hjortes Fitness Og Drægtighed. Videnskabelig Artikel 2013.Ulv i DanmarkNo ratings yet
- Undersøgelse Af Skab Hos Ulve I Spanien (Videnskabelig Artikel 2011)Document12 pagesUndersøgelse Af Skab Hos Ulve I Spanien (Videnskabelig Artikel 2011)Ulv i DanmarkNo ratings yet
- Artikel Om Brug Af Hyrdehunde Til Beskyttelse Af Husdyr GenereltDocument7 pagesArtikel Om Brug Af Hyrdehunde Til Beskyttelse Af Husdyr GenereltUlv i DanmarkNo ratings yet
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- Room For Wolf Comeback in The Netherlands: Rapport Om Ulvens Muligheder I HollandDocument23 pagesRoom For Wolf Comeback in The Netherlands: Rapport Om Ulvens Muligheder I HollandUlv i DanmarkNo ratings yet
- Rapport Om Status Af Og Trusler Imod Ulven I EuropaDocument27 pagesRapport Om Status Af Og Trusler Imod Ulven I EuropaUlv i DanmarkNo ratings yet
- Artikel Om Brug Af en Særlig Slags Elektrisk Hegn ("Electrified Fladry") Til Beskyttelse Af Husdyr Mod UlveDocument7 pagesArtikel Om Brug Af en Særlig Slags Elektrisk Hegn ("Electrified Fladry") Til Beskyttelse Af Husdyr Mod UlveUlv i DanmarkNo ratings yet
- Action Plan For The Conservation of The Wolves in EuropeDocument84 pagesAction Plan For The Conservation of The Wolves in EuropeUlv i DanmarkNo ratings yet
- Rapport Over Ulveangreb På Mennesker I Skandinavien de Sidste 300 ÅrDocument7 pagesRapport Over Ulveangreb På Mennesker I Skandinavien de Sidste 300 ÅrUlv i DanmarkNo ratings yet
- A History of Jackson County, OhioDocument202 pagesA History of Jackson County, Ohiovwindrow7070No ratings yet
- Chapter - 9 6 Class Science: The Living Organisms and Their SurroundingsDocument69 pagesChapter - 9 6 Class Science: The Living Organisms and Their SurroundingsShubham RajputNo ratings yet
- Rain of Gold by Victor VillaseñorDocument505 pagesRain of Gold by Victor VillaseñorArte Público Press69% (16)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (119)
- UntitledDocument2 pagesUntitledAisyahNo ratings yet
- Amateur Photographer - 21 November 2015Document84 pagesAmateur Photographer - 21 November 2015stamenkovskibNo ratings yet
- Sexism 001Document9 pagesSexism 001Rachelle GuevarraNo ratings yet
- Transfer Project Book Nº1 Screen SizeDocument30 pagesTransfer Project Book Nº1 Screen SizeEdgar EndressNo ratings yet
- Animal Skit (For Grades 2 & 3) : Scene 1: Animals and Their Unique Body PartsDocument12 pagesAnimal Skit (For Grades 2 & 3) : Scene 1: Animals and Their Unique Body PartsMarie Antoinette RoxasNo ratings yet
- DruiddDocument151 pagesDruiddRodney Mackay100% (3)
- Custer State Park - Guide To (2013) - 32 PagesDocument32 pagesCuster State Park - Guide To (2013) - 32 Pages88888888GZ88888888No ratings yet
- University of St. Thomas (Center For Irish Studies)Document20 pagesUniversity of St. Thomas (Center For Irish Studies)Martina SandrićNo ratings yet
- Comb MvbfbakingDocument12 pagesComb MvbfbakingCallum CurninNo ratings yet
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- Deer Predation vs StarvationDocument2 pagesDeer Predation vs StarvationIan BurtNo ratings yet
- Breaking Legacies by Zoe ReedDocument438 pagesBreaking Legacies by Zoe ReedNina Skromak100% (3)
- UntitledDocument7 pagesUntitledapi-233604231No ratings yet
- Baby Animals PDFDocument36 pagesBaby Animals PDFNga PhạmNo ratings yet
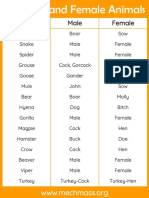
- Male and Female Animal NamesDocument7 pagesMale and Female Animal NamesMayur VaghelaNo ratings yet
- Preparing For Aptitude Tests: Career SkillsDocument39 pagesPreparing For Aptitude Tests: Career SkillsMalik Sajid SanwalNo ratings yet
- 2020-21 Big Game Status BookDocument200 pages2020-21 Big Game Status BookAaron Meier0% (1)
- Michigan Deer Harvest Report 2020Document77 pagesMichigan Deer Harvest Report 2020Arielle BreenNo ratings yet
- Kelompok 5 - Pengamatan Rusa Dengan Metode Scan SamplingDocument9 pagesKelompok 5 - Pengamatan Rusa Dengan Metode Scan SamplingXXX PegasuSNo ratings yet
- A Waldorf Reader For Early Third GradeDocument56 pagesA Waldorf Reader For Early Third GradeMagali De la Hoz100% (1)
- Game Animals of India 0 LydeDocument454 pagesGame Animals of India 0 Lydeshree2101No ratings yet
- Pearl A Grandmothers StoriesDocument136 pagesPearl A Grandmothers StoriesGary Loeb100% (2)
- Aptitude Tests Question Types and Scoring: Sample Numerical Aptitude Test QuestionsDocument34 pagesAptitude Tests Question Types and Scoring: Sample Numerical Aptitude Test QuestionsApoorv NigamNo ratings yet
- Big Game Guide: - Antelope - Bighorn Sheep - Deer - ElkDocument46 pagesBig Game Guide: - Antelope - Bighorn Sheep - Deer - ElkRoeHuntingResourcesNo ratings yet
- Ujian Ogos Bahasa Inggeris Tahun 3 Penulisan 2018Document14 pagesUjian Ogos Bahasa Inggeris Tahun 3 Penulisan 2018Kanagaa ManickamNo ratings yet
- Little Deer and The Tra1Document5 pagesLittle Deer and The Tra1jefferyNo ratings yet
- The Coordinative Way of Thinking in Eastern CulturesDocument21 pagesThe Coordinative Way of Thinking in Eastern CulturesmivanmegNo ratings yet
- Cave Art in EuropeDocument4 pagesCave Art in EuropeSahil PatelNo ratings yet
- Dog Training Journeys: A Guide to Training and Bonding with Your Mix-Breed DogFrom EverandDog Training Journeys: A Guide to Training and Bonding with Your Mix-Breed DogRating: 4.5 out of 5 stars4.5/5 (77)
- Welcome to the United States of Anxiety: Observations from a Reforming NeuroticFrom EverandWelcome to the United States of Anxiety: Observations from a Reforming NeuroticRating: 3.5 out of 5 stars3.5/5 (10)
- Mastering Parrot Behavior: A Step-by-Step Guide to Building a Strong Relationship with Your Avian FriendFrom EverandMastering Parrot Behavior: A Step-by-Step Guide to Building a Strong Relationship with Your Avian FriendRating: 4.5 out of 5 stars4.5/5 (69)
- Horse Training 101: Key Techniques for Every Horse OwnerFrom EverandHorse Training 101: Key Techniques for Every Horse OwnerRating: 4.5 out of 5 stars4.5/5 (27)
- The Asshole Survival Guide: How to Deal with People Who Treat You Like DirtFrom EverandThe Asshole Survival Guide: How to Deal with People Who Treat You Like DirtRating: 4 out of 5 stars4/5 (60)
- Cats Can Learn Too: A Simple Guide to Training Your Furry FriendFrom EverandCats Can Learn Too: A Simple Guide to Training Your Furry FriendRating: 4.5 out of 5 stars4.5/5 (55)