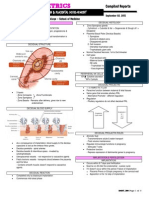
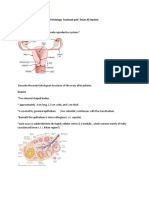
You might also like
- ACFrOgCu W0g9MxVK8YkEvfsqyGhjSstpPm3j2uJhpy4G69TRnw18zCsRC69oQlKhanJWV 6usc VqeS933XuRqaxNoOkxjXkvFwDAm6It15dFc0KjwYag EzHDH6St2KFBAy0eOIbDC WNeC9lpDocument18 pagesACFrOgCu W0g9MxVK8YkEvfsqyGhjSstpPm3j2uJhpy4G69TRnw18zCsRC69oQlKhanJWV 6usc VqeS933XuRqaxNoOkxjXkvFwDAm6It15dFc0KjwYag EzHDH6St2KFBAy0eOIbDC WNeC9lpUrvashi KhadeNo ratings yet
- Information Networks in The Mammary Gland: Lothar Hennighausen and Gertraud W. RobinsonDocument11 pagesInformation Networks in The Mammary Gland: Lothar Hennighausen and Gertraud W. RobinsonSueños De LunaNo ratings yet
- Molecular Mechanism of ImplantationDocument19 pagesMolecular Mechanism of ImplantationAbunu TilayeNo ratings yet
- Reproductive Biology and Technology in Animals PDFDocument126 pagesReproductive Biology and Technology in Animals PDFJuan CaroNo ratings yet
- Lactation PhysiologyDocument17 pagesLactation PhysiologysanathNo ratings yet
- Module 5Document10 pagesModule 5Phan MhiveNo ratings yet
- Breast Development PDFDocument26 pagesBreast Development PDFgiant nitaNo ratings yet
- EMBRYOLOGYDocument30 pagesEMBRYOLOGYMSc. PreviousNo ratings yet
- Buhimschi 2004 Endocrinology of LactationDocument17 pagesBuhimschi 2004 Endocrinology of LactationCarolina GómezNo ratings yet
- Oophorectomy &its Rolrole in Breast Cancer Risk ReductionDocument23 pagesOophorectomy &its Rolrole in Breast Cancer Risk ReductionNorhan Ahmed Ahmed Hassan El-MeniawyNo ratings yet
- Grimm 2020Document16 pagesGrimm 2020MD LarasatiNo ratings yet
- The Growing Fetus FertilizationDocument6 pagesThe Growing Fetus FertilizationJustJ ThingsNo ratings yet
- Implantation and Development of The PlacentaDocument8 pagesImplantation and Development of The PlacentaZulfi Nur Amrina RosyadaNo ratings yet
- Signalling Pathways Implicated in Early Mammary Gland Morphogenesis and Breast CancerDocument19 pagesSignalling Pathways Implicated in Early Mammary Gland Morphogenesis and Breast CancerhajimehikariNo ratings yet
- Fisiologia de La Secrecion Leche Materna Truchet2017Document45 pagesFisiologia de La Secrecion Leche Materna Truchet2017vanetoledoNo ratings yet
- Tutorial 4 Biology 2 Group 3 HN2Document3 pagesTutorial 4 Biology 2 Group 3 HN2Firdaus ZulkifliNo ratings yet
- CH 3Document8 pagesCH 3api-288494741No ratings yet
- GametogenesisDocument8 pagesGametogenesisSOPHIA ALESNANo ratings yet
- CVM Pres - IIDocument152 pagesCVM Pres - IIseraphim8No ratings yet
- 1.1c Implantation & Placental DevelopmentDocument5 pages1.1c Implantation & Placental DevelopmentBea Samonte100% (1)
- Department of Morphology: Penza State University Medical InstituteDocument18 pagesDepartment of Morphology: Penza State University Medical InstituteunnatiNo ratings yet
- Reproduction in AnimalsDocument12 pagesReproduction in AnimalsYuh moddaNo ratings yet
- Anatomy and Physiology GCSDocument7 pagesAnatomy and Physiology GCSSherry Ann FayeNo ratings yet
- Chapter - 3 Human ReproductionDocument3 pagesChapter - 3 Human ReproductionMr.Prashant SharmaNo ratings yet
- Normal PregnancyDocument65 pagesNormal PregnancyRhesie Joyce AguilarNo ratings yet
- Approach To Breast FeedingDocument13 pagesApproach To Breast FeedingsmokaNo ratings yet
- Obstetrics V10 Common Obstetric Conditions Chapter Pathology of The Placenta 1663285373Document11 pagesObstetrics V10 Common Obstetric Conditions Chapter Pathology of The Placenta 1663285373Nurliana NingsihNo ratings yet
- Mammary GlandDocument11 pagesMammary GlandPRITAM BHOIRNo ratings yet
- BlastulaDocument4 pagesBlastulaCyanDesNo ratings yet
- Why Is Placentation Abnormal in Preeclampsia?Document8 pagesWhy Is Placentation Abnormal in Preeclampsia?cecilliacynthiaNo ratings yet
- Postimplantation Development in MammalianDocument52 pagesPostimplantation Development in MammalianBiology BảoNo ratings yet
- 4 1Document265 pages4 1sillypoloNo ratings yet
- Fertilization Reflection PaperDocument2 pagesFertilization Reflection PaperCrisandro Allen Lazo100% (2)
- Abnormal ImplantationDocument3 pagesAbnormal ImplantationMuhammet Fatih CantepeNo ratings yet
- Last Minute Embryology: Human embryology made easy and digestible for medical and nursing studentsFrom EverandLast Minute Embryology: Human embryology made easy and digestible for medical and nursing studentsNo ratings yet
- Module 2 HandsoutDocument10 pagesModule 2 HandsoutErin SaavedraNo ratings yet
- Gametogenesis and FertilizationDocument10 pagesGametogenesis and FertilizationJustus MutuaNo ratings yet
- Obstetrics: The Normal Menstrual CycleDocument4 pagesObstetrics: The Normal Menstrual Cycleapi-3829364No ratings yet
- UntitledDocument234 pagesUntitledMarco Antonio AlvarezNo ratings yet
- Eterinary Bstetrics: - Dr. S.BalasubramanianDocument413 pagesEterinary Bstetrics: - Dr. S.BalasubramanianprernaNo ratings yet
- Phases of Embryonic DevelopmentDocument9 pagesPhases of Embryonic DevelopmentIshu ChoudharyNo ratings yet
- Prenatal GrowthDocument56 pagesPrenatal GrowthnavjotsinghjassalNo ratings yet
- Wella S, VIRINA Bsn-I: Review Question Reproductive SystemDocument5 pagesWella S, VIRINA Bsn-I: Review Question Reproductive SystemEllee HadesNo ratings yet
- Histo FemDocument9 pagesHisto FemzaidNo ratings yet
- CH 5 Implantation, Placental Dev (Part1)Document36 pagesCH 5 Implantation, Placental Dev (Part1)Tengku Chairannisa PutriNo ratings yet
- Embryology GastrulationDocument9 pagesEmbryology GastrulationRivera Gómez América LucíaNo ratings yet
- Regulation of Blastocyst Formation.Document24 pagesRegulation of Blastocyst Formation.christopher LopezNo ratings yet
- Embryology of Genital System & Congenital Anomalies: Kenbon S. (MS) January 30,2021Document56 pagesEmbryology of Genital System & Congenital Anomalies: Kenbon S. (MS) January 30,2021gimNo ratings yet
- 1st, 2nd & 3rdweeksDocument9 pages1st, 2nd & 3rdweeksPristine KrNo ratings yet
- Reproduction (M, F) : Lecturer: Dr. R. Ahangari University of Central Florida, OrlandoDocument10 pagesReproduction (M, F) : Lecturer: Dr. R. Ahangari University of Central Florida, Orlandokuku411No ratings yet
- Chapter 29 - Development and Inheritance: ST ND RD STDocument5 pagesChapter 29 - Development and Inheritance: ST ND RD STtomorrow.today.yesterday .yesterdayNo ratings yet
- Preclinical Anatomy Review 2023: For USMLE Step 1 and COMLEX-USA Level 1From EverandPreclinical Anatomy Review 2023: For USMLE Step 1 and COMLEX-USA Level 1Rating: 5 out of 5 stars5/5 (2)
- Naim LBM 1-1Document38 pagesNaim LBM 1-1dimasNo ratings yet
- EmbryologyDocument101 pagesEmbryologytawandarukwavamadisonNo ratings yet
- Funcion Endocrina de La Placenta HumanaDocument30 pagesFuncion Endocrina de La Placenta HumanaSofia RossoNo ratings yet
- USMLE NotesDocument165 pagesUSMLE NotesHerliani HalimNo ratings yet
- Sexual Reproduction and Meiosis: Four Haploid CellsDocument10 pagesSexual Reproduction and Meiosis: Four Haploid CellsJohnieer Bassem MoferdNo ratings yet
- LAB EXERCISE 2 - OogenesisDocument4 pagesLAB EXERCISE 2 - OogenesisGerald Angelo DeguinioNo ratings yet
- Reproduction PPT 6 PHYSIOLOGY OF PREGNANCYDocument98 pagesReproduction PPT 6 PHYSIOLOGY OF PREGNANCYlisanames.23No ratings yet
- Review of Fetal DevelopmentDocument71 pagesReview of Fetal DevelopmentlisafelixNo ratings yet
- NarasimhaDocument11 pagesNarasimhadaitya99No ratings yet
- VamanaDocument3 pagesVamanadaitya99No ratings yet
- VarahaDocument5 pagesVarahadaitya99No ratings yet
- ValmikiDocument2 pagesValmikidaitya99No ratings yet
- BrahmaDocument5 pagesBrahmadaitya99No ratings yet
- NaradDocument2 pagesNaraddaitya99No ratings yet
- Old World MonkeyDocument6 pagesOld World Monkeydaitya99No ratings yet
- Kingdom of BhutanDocument21 pagesKingdom of Bhutandaitya99No ratings yet
- Mammal: MammalsDocument13 pagesMammal: Mammalsdaitya99No ratings yet
- PrimatesDocument15 pagesPrimatesdaitya99No ratings yet
- Republic of Fiji: Navigation Search Fiji (Disambiguation)Document18 pagesRepublic of Fiji: Navigation Search Fiji (Disambiguation)daitya99No ratings yet
- List of Fossil PrimatesDocument20 pagesList of Fossil Primatesdaitya99No ratings yet
- Standard Assessment Form For Postgraduate Courses (Microbiology)Document15 pagesStandard Assessment Form For Postgraduate Courses (Microbiology)drunken monkeyNo ratings yet
- Suicide CaseDocument2 pagesSuicide Casedaitya99No ratings yet
- Ratna Shastra - The Mystery Hidden in GemstonesDocument35 pagesRatna Shastra - The Mystery Hidden in Gemstonesdaitya9957% (7)
- Marketing Study of Mango JuiceDocument18 pagesMarketing Study of Mango JuiceVijay ArapathNo ratings yet
- DR K.M.NAIR - GEOSCIENTIST EXEMPLARDocument4 pagesDR K.M.NAIR - GEOSCIENTIST EXEMPLARDrThrivikramji KythNo ratings yet
- Test On QuantifiersDocument1 pageTest On Quantifiersvassoula35No ratings yet
- 220hp Caterpillar 3306 Gardner Denver SSP Screw Compressor DrawingsDocument34 pages220hp Caterpillar 3306 Gardner Denver SSP Screw Compressor DrawingsJVMNo ratings yet
- Powerful Communication Tools For Successful Acupuncture PracticeDocument4 pagesPowerful Communication Tools For Successful Acupuncture Practicebinglei chenNo ratings yet
- Atlas of Feline Anatomy For VeterinariansDocument275 pagesAtlas of Feline Anatomy For VeterinariansДибензол Ксазепин100% (4)
- Chap 6 - Karen HorneyDocument95 pagesChap 6 - Karen HorneyDiana San JuanNo ratings yet
- ContinueDocument2 pagesContinueNeal ReppNo ratings yet
- Issue of HomosexualityDocument4 pagesIssue of HomosexualityT-2000No ratings yet
- Hodgkin LymphomaDocument44 pagesHodgkin LymphomaisnineNo ratings yet
- Pressure Classes: Ductile Iron PipeDocument4 pagesPressure Classes: Ductile Iron PipesmithNo ratings yet
- Index Medicus PDFDocument284 pagesIndex Medicus PDFVania Sitorus100% (1)
- TherabandDocument1 pageTherabandsuviacesoNo ratings yet
- Varioklav Steam Sterilizer 75 S - 135 S Technical SpecificationsDocument10 pagesVarioklav Steam Sterilizer 75 S - 135 S Technical Specificationssagor sagorNo ratings yet
- A Cook's Journey To Japan - Fish Tales and Rice Paddies 100 Homestyle Recipes From Japanese KitchensDocument306 pagesA Cook's Journey To Japan - Fish Tales and Rice Paddies 100 Homestyle Recipes From Japanese KitchensEthan F.100% (1)
- CFPB Discount Points Guidence PDFDocument3 pagesCFPB Discount Points Guidence PDFdzabranNo ratings yet
- ClistDocument14 pagesClistGuerraNo ratings yet
- Vaccination Schedule in Dogs and CatsDocument3 pagesVaccination Schedule in Dogs and CatsAKASH ANANDNo ratings yet
- Borelog CP.101Document1 pageBorelog CP.101radixkusumaNo ratings yet
- Intentions and Results ASFA and Incarcerated ParentsDocument10 pagesIntentions and Results ASFA and Incarcerated Parentsaflee123No ratings yet
- Cyber Safety PP Presentation For Class 11Document16 pagesCyber Safety PP Presentation For Class 11WAZ CHANNEL100% (1)
- Facts About Concussion and Brain Injury: Where To Get HelpDocument20 pagesFacts About Concussion and Brain Injury: Where To Get HelpJess GracaNo ratings yet
- Geography - Development (Rural - Urban Settlement)Document32 pagesGeography - Development (Rural - Urban Settlement)jasmine le rouxNo ratings yet
- Chi - Square Test: PG Students: DR Amit Gujarathi DR Naresh GillDocument32 pagesChi - Square Test: PG Students: DR Amit Gujarathi DR Naresh GillNaresh GillNo ratings yet
- Gay Costa Del Sol - 2010Document2 pagesGay Costa Del Sol - 2010gayinfospainNo ratings yet
- Cemco T80Document140 pagesCemco T80Eduardo Ariel Bernal100% (3)
- Social Style InventoryDocument12 pagesSocial Style InventoryMaheshwari JaniNo ratings yet
- 7 Fuel Failure in Water Reactors - Causes and MitigationDocument165 pages7 Fuel Failure in Water Reactors - Causes and MitigationLauri RubertiNo ratings yet
- Waste Heat Recovery UnitDocument15 pagesWaste Heat Recovery UnitEDUARDONo ratings yet
- Week5 6 2Document2 pagesWeek5 6 2SAMANIEGO BERMEO DAVID SEBASTIANNo ratings yet