You might also like
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (895)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- Bimodal Volcanism of The High Lava Plains and NW Basin and Range Age Progressive RhyolitesDocument22 pagesBimodal Volcanism of The High Lava Plains and NW Basin and Range Age Progressive RhyolitesreaderwriteNo ratings yet
- Fifty Years Ago: The Vaiont NightDocument4 pagesFifty Years Ago: The Vaiont NightreaderwriteNo ratings yet
- Carlins Fluid Flow BrevityDocument28 pagesCarlins Fluid Flow BrevityreaderwriteNo ratings yet
- G M S M D: Eology and Ineral Ystems of The IKE EpositDocument25 pagesG M S M D: Eology and Ineral Ystems of The IKE EpositreaderwriteNo ratings yet
- Opposing Mechanisms Support The Voluntary Forgetting of Unwanted MemoriesDocument11 pagesOpposing Mechanisms Support The Voluntary Forgetting of Unwanted MemoriesreaderwriteNo ratings yet
- Instructions For Building A Vita StoveDocument7 pagesInstructions For Building A Vita StovereaderwriteNo ratings yet
- Building The Harry Davis Single Shaft Vacuum DeAiring PugmillDocument10 pagesBuilding The Harry Davis Single Shaft Vacuum DeAiring PugmillreaderwriteNo ratings yet
- LimewashDocument11 pagesLimewashreaderwriteNo ratings yet
- Furnace, Gas-Fired SmeltingDocument4 pagesFurnace, Gas-Fired SmeltingJim100% (3)
- Making Building Blocks With The CINVA Ram Block PressDocument24 pagesMaking Building Blocks With The CINVA Ram Block Pressreaderwrite100% (1)
- Adobe NotesDocument24 pagesAdobe NotesreaderwriteNo ratings yet
- Neurobiology of EmpathyDocument33 pagesNeurobiology of EmpathyreaderwriteNo ratings yet
- 04 5792Document143 pages04 5792maouininNo ratings yet
- CINVA RAM For Making Adobe Building Blocks 1999Document26 pagesCINVA RAM For Making Adobe Building Blocks 1999readerwrite100% (1)
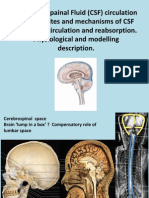
- Physics of Cerebrospinal Fluid CirculationDocument37 pagesPhysics of Cerebrospinal Fluid CirculationreaderwriteNo ratings yet
- Modern Adobe in New MexicoDocument29 pagesModern Adobe in New MexicoreaderwriteNo ratings yet
- Thallium IcpDocument9 pagesThallium IcpreaderwriteNo ratings yet
- The Impact of Neglect On The Developing Brain: Nature and NurtureDocument22 pagesThe Impact of Neglect On The Developing Brain: Nature and NurtureBrDuncPer100% (1)
- The Impact of Neglect On The Developing Brain: Nature and NurtureDocument22 pagesThe Impact of Neglect On The Developing Brain: Nature and NurtureBrDuncPer100% (1)
- Lifetime Victimization Children Mental HealthDocument15 pagesLifetime Victimization Children Mental HealthreaderwriteNo ratings yet
- Mindfulness Training and Neural Integration: Differentiation of Distinct Streams of Awareness and The Cultivation of Well BeingDocument9 pagesMindfulness Training and Neural Integration: Differentiation of Distinct Streams of Awareness and The Cultivation of Well BeingreaderwriteNo ratings yet
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5794)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (400)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (588)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (74)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (344)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1090)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2259)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (121)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- 72 Inventory Management and Financial PeDocument22 pages72 Inventory Management and Financial PeKiersteen AnasNo ratings yet
- Chapter 1 RHODADocument55 pagesChapter 1 RHODAJosenia ConstantinoNo ratings yet
- The Influence of Servant Leadership and Work Environment On Employee Happiness With Motivation As A Variable Intervening at PT PNM Pematang Siantar BranchDocument7 pagesThe Influence of Servant Leadership and Work Environment On Employee Happiness With Motivation As A Variable Intervening at PT PNM Pematang Siantar BranchInternational Journal of Innovative Science and Research TechnologyNo ratings yet
- AR Model Session2 Output: Install - Packages ("Forecast")Document30 pagesAR Model Session2 Output: Install - Packages ("Forecast")karthikmaddula007_66No ratings yet
- Statistics Chapter 10-12Document11 pagesStatistics Chapter 10-12Alna Gamulo LosaNo ratings yet
- Factors Affecting Employees Performance and Retent PDFDocument32 pagesFactors Affecting Employees Performance and Retent PDFUroOj SaleEmNo ratings yet
- CUHK STAT5102 Ch2Document25 pagesCUHK STAT5102 Ch2Tsoi Yun PuiNo ratings yet
- Marketing of Organic Food Products at Madurai City: P Shandini and N RamaniDocument21 pagesMarketing of Organic Food Products at Madurai City: P Shandini and N RamanishandiniNo ratings yet
- British Journal of Marketing Studies Vol.2, No.6, pp.52-67, October 2014 Published by European Centre For Research Training and Development UKDocument16 pagesBritish Journal of Marketing Studies Vol.2, No.6, pp.52-67, October 2014 Published by European Centre For Research Training and Development UKAbay ShiferaNo ratings yet
- 2020 Mock Exam A - Afternoon Session (With Solutions)Document62 pages2020 Mock Exam A - Afternoon Session (With Solutions)NikNo ratings yet
- A Study On Employability Skills of Mba Students From The Management Institutes in The State of Kerala Bindhu Ann Thomas & Dr. K. V. UnninarayananDocument7 pagesA Study On Employability Skills of Mba Students From The Management Institutes in The State of Kerala Bindhu Ann Thomas & Dr. K. V. UnninarayananBindhu Ann ThomasNo ratings yet
- Test of Hypothesis Involving MeansDocument12 pagesTest of Hypothesis Involving MeansRiski Andhika R, S. PdNo ratings yet
- The Process Capability AnalysisDocument18 pagesThe Process Capability AnalysisMohamed HamdyNo ratings yet
- Methods of Performance Analysis in Team Invasion Sports: A Systematic ReviewDocument13 pagesMethods of Performance Analysis in Team Invasion Sports: A Systematic ReviewGabriel AnjosNo ratings yet
- 1330 1507200472 PDFDocument17 pages1330 1507200472 PDFRosemarie SulibagaNo ratings yet
- Acceptability of Pan de YaconDocument11 pagesAcceptability of Pan de YaconJean CristobalNo ratings yet
- CH 09Document70 pagesCH 09Saied Aly SalamahNo ratings yet
- WWW Statisticshowto Com How To Do A T Test in Excel PDFDocument10 pagesWWW Statisticshowto Com How To Do A T Test in Excel PDFkokoyNo ratings yet
- Influence of School Proprietorship On The Achievement of Students Taught Oral English With Games TechniqueDocument7 pagesInfluence of School Proprietorship On The Achievement of Students Taught Oral English With Games TechniqueEditor IJTSRDNo ratings yet
- Impact of Training Development On The PerformanceDocument8 pagesImpact of Training Development On The PerformanceNawid AhmadiNo ratings yet
- Pan African University: Institute For Basic Sciences, Technology and InnovationDocument9 pagesPan African University: Institute For Basic Sciences, Technology and InnovationMarc MarinNo ratings yet
- U09d1 ANOVA F Stat Research QuestionDocument2 pagesU09d1 ANOVA F Stat Research Questionarhodes777No ratings yet
- Statistical PDFDocument10 pagesStatistical PDFRanchoNo ratings yet
- Sophomore ResearchDocument37 pagesSophomore Researchapi-501281962No ratings yet
- Stat 166 Final PaperDocument59 pagesStat 166 Final PaperElyk ZeuqsavNo ratings yet
- Del Rosario, A - Section B - Exelcise 4Document14 pagesDel Rosario, A - Section B - Exelcise 4ariane galeno100% (1)
- Improvement of Writing Skills Using Acti PDFDocument11 pagesImprovement of Writing Skills Using Acti PDFAhmadNo ratings yet
- Lab7 Panugao, KAC Th4 7Document3 pagesLab7 Panugao, KAC Th4 7Vanessa May BlancioNo ratings yet
- Workshop Calculos Bioindicadores PDFDocument66 pagesWorkshop Calculos Bioindicadores PDFOEF_1399No ratings yet
- A Critical Study of Effectiveness of Online Learning On Students' AchievementDocument7 pagesA Critical Study of Effectiveness of Online Learning On Students' AchievementJessica AprilyaniNo ratings yet