You might also like
- The Obesity Code - Summarized for Busy People: Unlocking the Secrets of Weight Loss: Based on the Book by Jason FungFrom EverandThe Obesity Code - Summarized for Busy People: Unlocking the Secrets of Weight Loss: Based on the Book by Jason FungNo ratings yet
- Eczema Relief - Ultimate Guide To Handle And Control Your EczemaFrom EverandEczema Relief - Ultimate Guide To Handle And Control Your EczemaNo ratings yet
- Greenfield BrochureDocument65 pagesGreenfield Brochurebalakumar8688No ratings yet
- The Bee SpaceDocument3 pagesThe Bee SpaceMalcolm SanfordNo ratings yet
- Solemnity of Saint Charles BorromeoDocument24 pagesSolemnity of Saint Charles BorromeoCharles Basti Garcia AlmonteNo ratings yet
- Guide To Intermittent Fasting: Learn The Most Fundamental Parts of This LifehackDocument25 pagesGuide To Intermittent Fasting: Learn The Most Fundamental Parts of This LifehackzvifridmanNo ratings yet
- Alzheimers Diet and SupplementsDocument5 pagesAlzheimers Diet and SupplementsRobert Edwards100% (1)
- The MIND Diet A Detailed Guide For Beginner1Document2 pagesThe MIND Diet A Detailed Guide For Beginner1GEMNo ratings yet
- Poli NationDocument25 pagesPoli NationrkvaitlaNo ratings yet
- Vitamins Promote Natural Disease ResistanceDocument2 pagesVitamins Promote Natural Disease ResistancelarrevityNo ratings yet
- Rocine 3Document55 pagesRocine 3Nevenka Blažik100% (1)
- Breakfast Fo Ods Satiety: Mary Ellen Camire & Megan BlackmoreDocument6 pagesBreakfast Fo Ods Satiety: Mary Ellen Camire & Megan BlackmoreDavid Dragoş100% (1)
- Insulin and Its Metabolic Effects Ron RosedaleDocument2 pagesInsulin and Its Metabolic Effects Ron RosedaleBuzoianu MariaNo ratings yet
- MSDS Lemongrass Oil NaturalDocument8 pagesMSDS Lemongrass Oil NaturalVeshal Purshothama SeelanNo ratings yet
- Empowered: a Cancer Guide to Healing, Surviving, and Feeling BetterFrom EverandEmpowered: a Cancer Guide to Healing, Surviving, and Feeling BetterNo ratings yet
- History of Ayurvedic MedicineDocument3 pagesHistory of Ayurvedic MedicineashahulhjNo ratings yet
- Coconut OilDocument2 pagesCoconut Oildimo100% (1)
- The 21-Day Immunity Plan: The Sunday Times Bestseller - 'A Perfect Way To Take The First Step To Transforming Your Life' - From The Foreword by Tom Watson - DR Aseem MalhotraDocument5 pagesThe 21-Day Immunity Plan: The Sunday Times Bestseller - 'A Perfect Way To Take The First Step To Transforming Your Life' - From The Foreword by Tom Watson - DR Aseem Malhotrawesyxaky0% (2)
- Benefits of Sun Flower SeedsDocument20 pagesBenefits of Sun Flower SeedsMohd Idris MohiuddinNo ratings yet
- Honeybee Lifespan Regulated by Vitellogenin ProteinDocument20 pagesHoneybee Lifespan Regulated by Vitellogenin ProteinPablo ACNo ratings yet
- Allocation Costs of Regeneration - Tail Regeneration Constrains Body Growth Under Low Food Availability in Juvenile LizardsDocument13 pagesAllocation Costs of Regeneration - Tail Regeneration Constrains Body Growth Under Low Food Availability in Juvenile LizardsKass PérezNo ratings yet
- The Effects of Larval Diet On Adult Life-History Traits of The Black Soldier Y, Hermetia Illucens (Diptera: Stratiomyidae)Document9 pagesThe Effects of Larval Diet On Adult Life-History Traits of The Black Soldier Y, Hermetia Illucens (Diptera: Stratiomyidae)Евгений ЯворскийNo ratings yet
- tmpF477 TMPDocument13 pagestmpF477 TMPFrontiersNo ratings yet
- Stynoski&O'Connell 2017Document6 pagesStynoski&O'Connell 2017catmyngiNo ratings yet
- Longevity, Fertility and FecunDocument10 pagesLongevity, Fertility and Fecunbwp7fdpr5qNo ratings yet
- Membrane Fluidity and The Ability of Domestic BirdDocument8 pagesMembrane Fluidity and The Ability of Domestic Birdsaifulmangopo123No ratings yet
- Altermatt 2010Document10 pagesAltermatt 2010Jéfferson J. Álvarez GómezNo ratings yet
- Fitness OECDocument13 pagesFitness OECC B MNo ratings yet
- 59haley Copeia PDFDocument9 pages59haley Copeia PDFPaula MoralesNo ratings yet
- Ontogeny of Large Birds: Migrants Do It Faster: S M Y Y - TDocument9 pagesOntogeny of Large Birds: Migrants Do It Faster: S M Y Y - TIván AlvaradoNo ratings yet
- Walters 2005Document21 pagesWalters 2005Ana AlfonsinNo ratings yet
- Bosholn Et Al. 2019Document12 pagesBosholn Et Al. 2019marianeNo ratings yet
- Hraniri Cu Polen Sau Fara PolenDocument14 pagesHraniri Cu Polen Sau Fara PolenCatalinNo ratings yet
- 53 Publication Snakes Survive StarvationDocument10 pages53 Publication Snakes Survive StarvationÑiýāž MāłïķNo ratings yet
- Control of Invertebrate Occupants of Nests: August 2015Document2 pagesControl of Invertebrate Occupants of Nests: August 2015MHS MEUTIA SOLEHAHNo ratings yet
- Tropea Et Al 2012 PDFDocument8 pagesTropea Et Al 2012 PDFAndrea Gabriela RuizNo ratings yet
- (Faza Baihaqi) Productive Performance and Blood Profiles of Laying Hens Fed HermetiaDocument8 pages(Faza Baihaqi) Productive Performance and Blood Profiles of Laying Hens Fed Hermetiafaza baihaqiNo ratings yet
- Egg Size, Eggshell Porosity, and Incubation Period in The Marine Bird Family AlcidaeDocument9 pagesEgg Size, Eggshell Porosity, and Incubation Period in The Marine Bird Family AlcidaeAyyach FatehNo ratings yet
- Phytase Addition in PF - 3Document24 pagesPhytase Addition in PF - 3Vishwanath HebbiNo ratings yet
- Thesis 0.8Document9 pagesThesis 0.8jocyeoNo ratings yet
- tmp84CE TMPDocument13 pagestmp84CE TMPFrontiersNo ratings yet
- Aging Cell - 2007 - Robert - Testing The Free Radical Theory of Aging Hypothesis Physiological Differences in Long LivedDocument10 pagesAging Cell - 2007 - Robert - Testing The Free Radical Theory of Aging Hypothesis Physiological Differences in Long LivedAndrea G.No ratings yet
- How Physical Beetle Traits Impact FossilizationDocument6 pagesHow Physical Beetle Traits Impact FossilizationJean-Christophe ChampagneNo ratings yet
- Carmona Et Al. 2013 - Acta OecologicaDocument7 pagesCarmona Et Al. 2013 - Acta OecologicaCarlos Pérez CarmonaNo ratings yet
- Gartner 2020Document10 pagesGartner 2020RICARDO AYALA PE�ANo ratings yet
- Metabolite Support of Long-Term Storage of Sperm in The Spermatheca of Honeybee (Apis Mellifera) QueensDocument14 pagesMetabolite Support of Long-Term Storage of Sperm in The Spermatheca of Honeybee (Apis Mellifera) QueensJuan Pedro CabezaNo ratings yet
- Effect of Migratory Habits in Behavioral, Physiological and Morphological Differentiation of VertebratesDocument8 pagesEffect of Migratory Habits in Behavioral, Physiological and Morphological Differentiation of VertebratesDaniel Bustos ForeroNo ratings yet
- Hamer Et Al 2006 Carrion V FruitDocument9 pagesHamer Et Al 2006 Carrion V FruitKellyta RodriguezNo ratings yet
- Role of Oxidative Stress in Drosophila AgingDocument13 pagesRole of Oxidative Stress in Drosophila Agingdmchoi87No ratings yet
- Desarrollo Embrionario PollosDocument7 pagesDesarrollo Embrionario PollosSharon TarquiniNo ratings yet
- Temperature and Seed Storage Longevity: J. B. Dickie, R. H. Ellisf, H. L. Kraakj, K. Ryder and P. B. TompsettDocument8 pagesTemperature and Seed Storage Longevity: J. B. Dickie, R. H. Ellisf, H. L. Kraakj, K. Ryder and P. B. Tompsettau2011auNo ratings yet
- PDF ProcRoySocLondgradientDocument8 pagesPDF ProcRoySocLondgradientmaguitcoolNo ratings yet
- Livestock Science: ArticleinfoDocument7 pagesLivestock Science: ArticleinfoAdín Daza GarateNo ratings yet
- A Description of The Potential Growth and Body Composition of Two Commercial Broiler StrainsDocument29 pagesA Description of The Potential Growth and Body Composition of Two Commercial Broiler StrainsFabianeNo ratings yet
- Adams Et Al. 2002Document7 pagesAdams Et Al. 2002Aleix Alburnà BuscatóNo ratings yet
- The Foraging Behaviour of Honey Bees, Apis Mellifera: A ReviewDocument11 pagesThe Foraging Behaviour of Honey Bees, Apis Mellifera: A ReviewWaqar HassanNo ratings yet
- Production of Sexuals and Mating Frequency in The Stingless Bee Tetragonisca Angustula (Latreille) (Hymenoptera, Apidae)Document9 pagesProduction of Sexuals and Mating Frequency in The Stingless Bee Tetragonisca Angustula (Latreille) (Hymenoptera, Apidae)GabrielNo ratings yet
- Baudouin Ecology of Snakes Paper - December 2023Document8 pagesBaudouin Ecology of Snakes Paper - December 2023benedict.callebautNo ratings yet
- Journal of Zoology - 2008 - Gartner - Adaptation in The African Egg Eating Snake A Comparative Approach To A Classic StudyDocument7 pagesJournal of Zoology - 2008 - Gartner - Adaptation in The African Egg Eating Snake A Comparative Approach To A Classic StudyMichael Van der MerweNo ratings yet
- Berthouly FuncEc2008Document10 pagesBerthouly FuncEc2008richnerNo ratings yet
- AteneoDocument8 pagesAteneofabiandionisioNo ratings yet
- APIPUNTURADocument6 pagesAPIPUNTURAfabiandionisioNo ratings yet
- 09 Residuos Mieles CubanasDocument5 pages09 Residuos Mieles CubanasfabiandionisioNo ratings yet
- Control Varroa MiteDocument6 pagesControl Varroa MitefabiandionisioNo ratings yet
- 2006 BeenomeDocument19 pages2006 BeenomefabiandionisioNo ratings yet
- 10 Residuos Control Legislacion UsaDocument0 pages10 Residuos Control Legislacion UsafabiandionisioNo ratings yet
- Flight and FightDocument12 pagesFlight and FightfabiandionisioNo ratings yet
- Abejas Cria OxavarDocument4 pagesAbejas Cria OxavarfabiandionisioNo ratings yet
- Colony Record Card - Master BeekeeperDocument1 pageColony Record Card - Master BeekeeperfabiandionisioNo ratings yet
- Moving BeesDocument2 pagesMoving BeesfabiandionisioNo ratings yet
- De Facto - Life Mel Is Effective!: Dr. Olga OlizkiDocument0 pagesDe Facto - Life Mel Is Effective!: Dr. Olga OlizkifabiandionisioNo ratings yet
- Varroa Cuadros ViejosDocument6 pagesVarroa Cuadros ViejosfabiandionisioNo ratings yet
- Benefits of Honey WoundsDocument8 pagesBenefits of Honey WoundsfabiandionisioNo ratings yet
- Apimondia 2007: 40th International Apicultural Congress in MelbourneDocument32 pagesApimondia 2007: 40th International Apicultural Congress in MelbournefabiandionisioNo ratings yet
- Honeys From MoliseDocument4 pagesHoneys From MolisefabiandionisioNo ratings yet
- Bactericidal Activity of Different HoneysDocument4 pagesBactericidal Activity of Different HoneysMuftihat IsrarNo ratings yet
- Cambios ColorDocument7 pagesCambios ColorfabiandionisioNo ratings yet
- Bee Venom CreamDocument2 pagesBee Venom CreamfabiandionisioNo ratings yet
- Efecto Olor en AbejasDocument11 pagesEfecto Olor en AbejasfabiandionisioNo ratings yet
- Idaho Bee Crop Feb 2006Document2 pagesIdaho Bee Crop Feb 2006fabiandionisioNo ratings yet
- WWF Press Release COCA COLADocument3 pagesWWF Press Release COCA COLAfabiandionisioNo ratings yet
- Microorganisms in Honey: A Review of Sources and Significance for Quality and SafetyDocument26 pagesMicroorganisms in Honey: A Review of Sources and Significance for Quality and SafetyfabiandionisioNo ratings yet
- White Coat HypertensionDocument10 pagesWhite Coat HypertensionfabiandionisioNo ratings yet
- Anticoncepc HormonalDocument10 pagesAnticoncepc HormonalfabiandionisioNo ratings yet
- Pseudo Ciano SisDocument2 pagesPseudo Ciano SisfabiandionisioNo ratings yet
- Honey CristallizationDocument3 pagesHoney CristallizationfabiandionisioNo ratings yet
- Pfizer Pays Record $2.3B Fine for Illegal Drug MarketingDocument2 pagesPfizer Pays Record $2.3B Fine for Illegal Drug MarketingfabiandionisioNo ratings yet
- Dos and DonTs in Web DesignDocument27 pagesDos and DonTs in Web DesignfabiandionisioNo ratings yet
- 10' Smoking CessationDocument1 page10' Smoking CessationfabiandionisioNo ratings yet
- Honeys From MoliseDocument4 pagesHoneys From MolisefabiandionisioNo ratings yet
- DepresijaDocument6 pagesDepresijaChristopher PhillipsNo ratings yet
- Laryngeal ObstructionDocument59 pagesLaryngeal ObstructionpravinNo ratings yet
- 08 DN 005 BA Int OkDocument64 pages08 DN 005 BA Int OkvbogachevNo ratings yet
- Platelet Rich Plasma in OrthopaedicsDocument269 pagesPlatelet Rich Plasma in OrthopaedicsBelinda Azhari SiswantoNo ratings yet
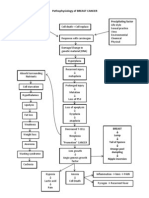
- Pathophysiology of BREAST CANCERDocument1 pagePathophysiology of BREAST CANCERAlinor Abubacar100% (6)
- Abdominal Incisions and Sutures in Obstetrics and GynaecologyDocument6 pagesAbdominal Incisions and Sutures in Obstetrics and GynaecologyFebyan AbotNo ratings yet
- Irritable Bowel Syndrome Natural SolutionsDocument25 pagesIrritable Bowel Syndrome Natural SolutionsBrettNo ratings yet
- Syrgery Mock 2Document8 pagesSyrgery Mock 2aa.No ratings yet
- Chapter 41 - Thoracic Outlet Syndrome SynonymsDocument8 pagesChapter 41 - Thoracic Outlet Syndrome SynonymsPiero Massafra100% (1)
- Properties of Skeletal MuscleDocument21 pagesProperties of Skeletal Musclenirilib72% (18)
- Application of Proper Draping-Final DemoDocument15 pagesApplication of Proper Draping-Final DemoMarife Hernandez Gelin80% (15)
- Kantor Cabang: BANJARMASIN - 1701 FKTP: Kertak Hanyar - 17040601Document6 pagesKantor Cabang: BANJARMASIN - 1701 FKTP: Kertak Hanyar - 17040601Ic-tika Siee ChuabbieNo ratings yet
- Chapter 3Document2 pagesChapter 3Rachel Marie M. Gania100% (1)
- CH 3 - Study GuideDocument13 pagesCH 3 - Study Guide2688gieNo ratings yet
- UQU SLE CORRECTED FILE by DR Samina FidaDocument537 pagesUQU SLE CORRECTED FILE by DR Samina Fidaasma .sassi100% (1)
- NURS1603 Course Outline PDFDocument19 pagesNURS1603 Course Outline PDFYip Ka YiNo ratings yet
- HemophiliaDocument27 pagesHemophiliaDorothy Pearl Loyola Palabrica100% (1)
- Neuromuscular Junction Blocking AgentsDocument9 pagesNeuromuscular Junction Blocking AgentsSyvNo ratings yet
- Delhi Public School Bangalore North Science Assignment KeyDocument3 pagesDelhi Public School Bangalore North Science Assignment KeyJanaki KrishnanNo ratings yet
- Complaint Affidavit SampleDocument13 pagesComplaint Affidavit SampleGertrude PillenaNo ratings yet
- 7/23/2016 Cristina S. Nebres Mindanao State University at Naawan 1Document32 pages7/23/2016 Cristina S. Nebres Mindanao State University at Naawan 1Laila UbandoNo ratings yet
- Anatomy & Physiology QuestionsDocument10 pagesAnatomy & Physiology Questionskrishna chandrakani100% (2)
- Unit 1 The Integumentary SystemDocument3 pagesUnit 1 The Integumentary SystemSharva BhasinNo ratings yet
- Traumatic Brain Injury PresentationDocument14 pagesTraumatic Brain Injury Presentationapi-413607178No ratings yet
- Abnormal heart sounds explainedDocument3 pagesAbnormal heart sounds explainedmuhammad azizulhakimNo ratings yet
- Advanced Trauma Life SupportDocument452 pagesAdvanced Trauma Life Supportnatalia100% (7)
- Physiology of The Liver: Corresponding AuthorDocument12 pagesPhysiology of The Liver: Corresponding AuthorMansour HazaNo ratings yet
- Pathologyofdiseasesof Geriatricexoticmammals: Drury R. Reavill,, Denise M. ImaiDocument34 pagesPathologyofdiseasesof Geriatricexoticmammals: Drury R. Reavill,, Denise M. ImaiRaquel MotaNo ratings yet
- Potassium Persulfate InformationDocument3 pagesPotassium Persulfate InformationmeimeiliuNo ratings yet
- Hypertension ControlDocument7 pagesHypertension ControlXyla CullenNo ratings yet