You might also like
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5794)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1090)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (895)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (588)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (344)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (121)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (400)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2259)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (74)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- Pages From 1894 Carey The Biochemic System of Medicine-Part 1of 9Document51 pagesPages From 1894 Carey The Biochemic System of Medicine-Part 1of 9Shiva Maru100% (3)
- Revision Practice Questions - Cells & TransportDocument11 pagesRevision Practice Questions - Cells & TransportMarina BrazendaleNo ratings yet
- Collision Between Ship and Whales 2001Document41 pagesCollision Between Ship and Whales 2001Vicky Concha AquevequeNo ratings yet
- Paper Nutrition and Fertility Dairy CowsDocument5 pagesPaper Nutrition and Fertility Dairy CowsVicky Concha AquevequeNo ratings yet
- Current Advances in Gestation and Parturition in Cats and DogsDocument12 pagesCurrent Advances in Gestation and Parturition in Cats and DogsVicky Concha AquevequeNo ratings yet
- Jensen, P. 2006Document13 pagesJensen, P. 2006Vicky Concha AquevequeNo ratings yet
- Antioxidantes en CaballosDocument6 pagesAntioxidantes en CaballosVicky Concha AquevequeNo ratings yet
- Model Answers: Chapter 3 Movement of Substances Across A Plasma MembraneDocument4 pagesModel Answers: Chapter 3 Movement of Substances Across A Plasma MembraneimanNo ratings yet
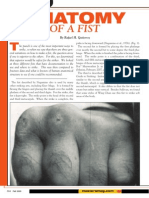
- Anatomy of FistDocument4 pagesAnatomy of FistCreative Flow Media100% (1)
- Aubf Prelim 1Document59 pagesAubf Prelim 1Kat JornadalNo ratings yet
- Ayurveda - Neurogenic BladderDocument3 pagesAyurveda - Neurogenic BladderyogasanaNo ratings yet
- Chemical Coordination and IntegrationDocument24 pagesChemical Coordination and IntegrationMahalaksshmi .DNo ratings yet
- Femoral EmbolectomyDocument13 pagesFemoral EmbolectomyAndrea Aji SanayaNo ratings yet
- SimpleNursing Study GuidesDocument5 pagesSimpleNursing Study Guidessilgabernard100% (1)
- Cell Structure and FunctionDocument33 pagesCell Structure and FunctionRonald DarwinNo ratings yet
- Connective Tissue NotesDocument6 pagesConnective Tissue Notesferdinand padillaNo ratings yet
- Peng2020 EFEK VITAMIN D PADA OBATDocument10 pagesPeng2020 EFEK VITAMIN D PADA OBATrifkiNo ratings yet
- Cabg 130102113345 Phpapp01Document29 pagesCabg 130102113345 Phpapp01md.dascalescu2486No ratings yet
- RH Incompatibility: Go ToDocument4 pagesRH Incompatibility: Go ToFadhila AnasNo ratings yet
- Science PMRDocument72 pagesScience PMRJiaYing WinNo ratings yet
- Mitosis and Meiosis Webquest 17Document3 pagesMitosis and Meiosis Webquest 17api-260817414No ratings yet
- IGCSE Biology Revision Exercise (Chap 5-8) Paper 2 1Document19 pagesIGCSE Biology Revision Exercise (Chap 5-8) Paper 2 1Sze Jack100% (1)
- PBL 12 - Adrenal GlandsDocument7 pagesPBL 12 - Adrenal GlandsKrishna OochitNo ratings yet
- Embryology Notes - emDocument18 pagesEmbryology Notes - emrigario80% (5)
- Arnold Jacobs: His Global Influence: Midwest Clinic, Thursday, December 17 12:00-1:00pmDocument17 pagesArnold Jacobs: His Global Influence: Midwest Clinic, Thursday, December 17 12:00-1:00pmAlex MoralesNo ratings yet
- Steps To Prepare The Divine Golden BookDocument2 pagesSteps To Prepare The Divine Golden Bookdesign12No ratings yet
- Sensory and Perceptual AlterationsDocument11 pagesSensory and Perceptual Alterationsbeverly_domingoNo ratings yet
- Irb DocumentsDocument17 pagesIrb Documentsapi-486275760No ratings yet
- AGE NCPDocument4 pagesAGE NCPXane Tañada DiazNo ratings yet
- Acute Dyspnoea and Haemoptysis My Notes Add Jimmy's PDF Notes On & OrganiseDocument171 pagesAcute Dyspnoea and Haemoptysis My Notes Add Jimmy's PDF Notes On & OrganisewwwrgrobinNo ratings yet
- Respiratory System - TransesDocument4 pagesRespiratory System - TransesMLS1C - PLARISAN, VALENT DAVENo ratings yet
- Untitled13 PDFDocument16 pagesUntitled13 PDFElizabeth LeonNo ratings yet
- 2011 - Mousavi Et Al - A General Overview On Manganese (MN) Importance For Crops ProductionDocument5 pages2011 - Mousavi Et Al - A General Overview On Manganese (MN) Importance For Crops ProductionHans Kristian AkarNo ratings yet
- Alchohol in Blood - UrineDocument19 pagesAlchohol in Blood - UrineDangLuuHaiNo ratings yet
- Immunopharmacology - IntroductionDocument51 pagesImmunopharmacology - IntroductionIsrat Jahan Surovy 2225400672No ratings yet