You might also like
- Kumo Desu Ga, Nani Ka - Volume 03 PDFDocument372 pagesKumo Desu Ga, Nani Ka - Volume 03 PDFMatia Gabriel100% (8)
- Business Spotlight July 2020 PDFDocument107 pagesBusiness Spotlight July 2020 PDFArie Ngasal100% (1)
- Social Impacts of Climate Change in Peru: A District Level Analysis of the Effects of Recent and Future Climate Change on Human Development and InequalityDocument36 pagesSocial Impacts of Climate Change in Peru: A District Level Analysis of the Effects of Recent and Future Climate Change on Human Development and InequalityCharly D WhiteNo ratings yet
- Understanding Market Channels and Alternatives For Commercial Catfish Farmers (PDFDrive)Document95 pagesUnderstanding Market Channels and Alternatives For Commercial Catfish Farmers (PDFDrive)AmiibahNo ratings yet
- The Global Seafood Revolution: George Chamberlain President Global Aquaculture AllianceDocument37 pagesThe Global Seafood Revolution: George Chamberlain President Global Aquaculture AllianceLuisNo ratings yet
- The Future of Securitization: 5 Annual Credit Risk ConferenceDocument13 pagesThe Future of Securitization: 5 Annual Credit Risk Conferenceanon-121346No ratings yet
- Interaction of Nanoclay-Reinforced (Esse Parece Ter Tudo o Que Eu Quero Fazer)Document24 pagesInteraction of Nanoclay-Reinforced (Esse Parece Ter Tudo o Que Eu Quero Fazer)ElesseaNo ratings yet
- GNN3 - M. Roco, NSF, USDocument52 pagesGNN3 - M. Roco, NSF, USMaterials Research InstituteNo ratings yet
- Prezentare WAGES 2007Document36 pagesPrezentare WAGES 2007Ionut DraganNo ratings yet
- The Philippine Debt Audit Campaign: A Renewed Call To The 14 Congress and The Filipino PeopleDocument24 pagesThe Philippine Debt Audit Campaign: A Renewed Call To The 14 Congress and The Filipino Peopletessafaye28No ratings yet
- Honjo Ts 1Document101 pagesHonjo Ts 1coachmaregaNo ratings yet
- Online Forum Portal for Human Flesh Search Discussions and ActivitiesDocument1 pageOnline Forum Portal for Human Flesh Search Discussions and ActivitiesCarlosNo ratings yet
- "Billionares": and They Were Not Happy About ItDocument36 pages"Billionares": and They Were Not Happy About Itkhanriyaz23941560No ratings yet
- River Pollution Incidents 2013Document17 pagesRiver Pollution Incidents 2013roshansanyasi50No ratings yet
- Operations & Supply Chain Management (Unit 3)Document31 pagesOperations & Supply Chain Management (Unit 3)Kiril IlievNo ratings yet
- 11_Sarkodie _& al_2016_on a_multivariate_analysis_with electricityDocument9 pages11_Sarkodie _& al_2016_on a_multivariate_analysis_with electricitykmlhectorseth94No ratings yet
- The Suitability of The Marine Biotic Index (AMBI) To New Impact Sources Along European CoastsDocument13 pagesThe Suitability of The Marine Biotic Index (AMBI) To New Impact Sources Along European CoastsJUAN PABLO PINO CATALANNo ratings yet
- Nickel Laterite DepositDocument23 pagesNickel Laterite DepositPutra100% (2)
- (Elias, 2002) Nickel Laterite Deposits - Geological Overview, Resources and ExploitationDocument24 pages(Elias, 2002) Nickel Laterite Deposits - Geological Overview, Resources and ExploitationRika yustika100% (1)
- Climate20Change20and20Waste20Management Chidozie20NnajiDocument19 pagesClimate20Change20and20Waste20Management Chidozie20NnajiChares Joke LangNo ratings yet
- A Review of Groundwater Issues, ReccomendationsDocument17 pagesA Review of Groundwater Issues, ReccomendationsAtami writeNo ratings yet
- NAPCCDocument31 pagesNAPCCPraveen Kumar GNo ratings yet
- Eco Background Final enDocument42 pagesEco Background Final enAurelian Buliga-StefanescuNo ratings yet
- Synthesis and Characterization of Fe-Doped Tio2 Photocatalyst by The Sol-Gel MethodDocument6 pagesSynthesis and Characterization of Fe-Doped Tio2 Photocatalyst by The Sol-Gel MethodsatishNo ratings yet
- B-019 Update: Phytoplankton, Robots, and FunDocument29 pagesB-019 Update: Phytoplankton, Robots, and FunRUCOOLNo ratings yet
- Seminar Teknik Untirta 2010Document23 pagesSeminar Teknik Untirta 2010Wawan GunawanNo ratings yet
- Ijaz Hussain PDFDocument25 pagesIjaz Hussain PDFsamashrafNo ratings yet
- Lecture 1Document10 pagesLecture 1shahzodrahmatillayev12No ratings yet
- 34 - CO2 Emissions in The USA New Insights Based On ANN ApproachDocument25 pages34 - CO2 Emissions in The USA New Insights Based On ANN ApproachTheRealCodiciaNo ratings yet
- Application of Indicator Kriging To Evaluate The Probability of Exceeding Nitrate Contamination ThresholdsDocument11 pagesApplication of Indicator Kriging To Evaluate The Probability of Exceeding Nitrate Contamination ThresholdsDayana Milagros Guerra NayraNo ratings yet
- Chap2 3Document46 pagesChap2 3Abhi KaleNo ratings yet
- Volcanic Ash, Metakaolin and Ordinary Portland Cement Blends in Concrete ProductionDocument4 pagesVolcanic Ash, Metakaolin and Ordinary Portland Cement Blends in Concrete ProductionJeriel PacerNo ratings yet
- Mauritius 2000-2006 National Greenhouse Gas Inventory ReportDocument297 pagesMauritius 2000-2006 National Greenhouse Gas Inventory ReportAshutosh Kumar SinghNo ratings yet
- Element Mobility Due To Hydrothermal Alteration in Los Azufres Geothermal Field, MexicoDocument5 pagesElement Mobility Due To Hydrothermal Alteration in Los Azufres Geothermal Field, MexicoSatryo BNo ratings yet
- Impact of Climate Change On Groundwater SystemDocument49 pagesImpact of Climate Change On Groundwater SystemMahmoud I. MahmoudNo ratings yet
- Determinación de La Respiración Del Suelo, Bajo El Uso de Diferentes XenobioticosDocument3 pagesDeterminación de La Respiración Del Suelo, Bajo El Uso de Diferentes Xenobioticoscristian najarNo ratings yet
- Energy Efficiency and Renewable Energy Sources in Latvia: Inguna Ozolina Latvian Investment and Development AgencyDocument22 pagesEnergy Efficiency and Renewable Energy Sources in Latvia: Inguna Ozolina Latvian Investment and Development AgencyEnergy dealersNo ratings yet
- HajeniusDocument28 pagesHajeniuslinh myNo ratings yet
- Goal: Climate ProtectionDocument9 pagesGoal: Climate ProtectionM-NCPPCNo ratings yet
- Changing Climate and Poverty in Vietnam: Searching For Pro-Poor Adaptation Policies - PresentationDocument23 pagesChanging Climate and Poverty in Vietnam: Searching For Pro-Poor Adaptation Policies - PresentationADB Poverty ReductionNo ratings yet
- Exb Practica 04Document2 pagesExb Practica 04Vinc Rivera GgamarNo ratings yet
- The Coffee Sector Presentation - KenyaDocument28 pagesThe Coffee Sector Presentation - KenyaDeepak Shankar0% (1)
- Common Ion On SlakingDocument4 pagesCommon Ion On SlakingProcess EngineerNo ratings yet
- ANNUAL REPORT 2011 RECORD-BREAKING CAPACITYDocument35 pagesANNUAL REPORT 2011 RECORD-BREAKING CAPACITYKathleen Kaye CastilloNo ratings yet
- Overview of U.S. Legislation and Regulations Affecting Offshore Natural Gas and Oil ActivityDocument20 pagesOverview of U.S. Legislation and Regulations Affecting Offshore Natural Gas and Oil ActivityAmir RashidiNo ratings yet
- Resources Reports Iceland-LabourMarketDevelopmentandPolicyDocument30 pagesResources Reports Iceland-LabourMarketDevelopmentandPolicyxoqvyxqipnffqjkypfNo ratings yet
- Ad MeDocument10 pagesAd MeRudhy Andry TanjungNo ratings yet
- Zero Waste Policy For Municipal Solid Waste in TaiwanoTHBDocument5 pagesZero Waste Policy For Municipal Solid Waste in TaiwanoTHBjoshlamNo ratings yet
- Sustainability and Energy ?Document26 pagesSustainability and Energy ?DaisyNo ratings yet
- ARR Mineral Sands 2017Document14 pagesARR Mineral Sands 2017Julio ArmandoNo ratings yet
- Listeriosis in The United States: Benjamin J. Silk, PHD, MPHDocument25 pagesListeriosis in The United States: Benjamin J. Silk, PHD, MPHmanuel camposNo ratings yet
- Overcapacity As A Barrier To Renewable Energy Depl PDFDocument11 pagesOvercapacity As A Barrier To Renewable Energy Depl PDFrichmondNo ratings yet
- Vol18Sabah Section2Document38 pagesVol18Sabah Section2Naziemi AhmadNo ratings yet
- Economic - Social - Performance 5 Yr (Eng)Document25 pagesEconomic - Social - Performance 5 Yr (Eng)Thaksin ShinawatraNo ratings yet
- Evidence of Global Warming: Hurricane and Temperature Data AnalysisDocument7 pagesEvidence of Global Warming: Hurricane and Temperature Data AnalysisJuan José Zapata FrancoNo ratings yet
- Effects of A Silica-Based Feed Supplement On Performance, Health, and Litter Quality of Growing TurkeysDocument7 pagesEffects of A Silica-Based Feed Supplement On Performance, Health, and Litter Quality of Growing TurkeysAJ ManurungNo ratings yet
- 105 Rainfall Characteristics During Pre - and Post-OnsetDocument9 pages105 Rainfall Characteristics During Pre - and Post-OnsetTeuku FerijalNo ratings yet
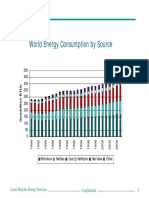
- World Energy Consumption by Source: Louis Dreyfus Energy ServicesDocument12 pagesWorld Energy Consumption by Source: Louis Dreyfus Energy ServicesARMANDO HERNANDEZNo ratings yet
- DSG20 Brief 20 EnglishDocument8 pagesDSG20 Brief 20 EnglishAng SHNo ratings yet
- Changes in Population ActivityDocument2 pagesChanges in Population ActivityDearbhla HubbardNo ratings yet
- Hydrogeochemistry Fundamentals and Advances, Environmental Analysis of GroundwaterFrom EverandHydrogeochemistry Fundamentals and Advances, Environmental Analysis of GroundwaterNo ratings yet
- Catfish: OsdcptjkDocument11 pagesCatfish: Osdcptjkenzo abrahamNo ratings yet
- SardinesDocument78 pagesSardinesGD MEPFCNo ratings yet
- Food and Feeding Habits of Fishes in ReservoirsDocument12 pagesFood and Feeding Habits of Fishes in ReservoirsFeroz KhanNo ratings yet
- Campbeltown's Quays, Coal and Sailing ShipsDocument4 pagesCampbeltown's Quays, Coal and Sailing ShipsKintyre On Record100% (1)
- Article1380270990 - Fadaeifard Et AlDocument5 pagesArticle1380270990 - Fadaeifard Et AlAdam PratamaNo ratings yet
- Ports&Harbors (List of Ports and Harbors in PH)Document8 pagesPorts&Harbors (List of Ports and Harbors in PH)Mark Barruga Duque100% (1)
- Manual t750 eDocument47 pagesManual t750 elordNo ratings yet
- Toba Lake StoryDocument3 pagesToba Lake Storyrakey alkautsarNo ratings yet
- CookbookDocument940 pagesCookbookChefe Jose Antonio OlimNo ratings yet
- Unit Test - Iii (Sample Paper) Class - Ukg Subject - English: NAME: - M.M.: 35Document15 pagesUnit Test - Iii (Sample Paper) Class - Ukg Subject - English: NAME: - M.M.: 35dipanwita samantaNo ratings yet
- Marine Ecosystem TypesDocument20 pagesMarine Ecosystem Typesfunky goodNo ratings yet
- Basic Fantasy Creature List by Mike Roop r3 De1Document101 pagesBasic Fantasy Creature List by Mike Roop r3 De1Gab G.No ratings yet
- Technology and Livelihood Education: Cookery NC IiDocument34 pagesTechnology and Livelihood Education: Cookery NC IiJocel AlicanteNo ratings yet
- Technology and Home EconomicsDocument8 pagesTechnology and Home EconomicsAngel Sealna100% (1)
- Types of Ships: Bulk Carriers, Oil Tankers and Container VesselsDocument36 pagesTypes of Ships: Bulk Carriers, Oil Tankers and Container VesselsAktarojjaman MiltonNo ratings yet
- The Impact of The Northern Snakehead On Aquatic Life in The UsDocument7 pagesThe Impact of The Northern Snakehead On Aquatic Life in The Usapi-620287053No ratings yet
- Project 2Document5 pagesProject 2khanhphuongdo156No ratings yet
- Adaptation and Survival: of Organisms in The Changing WorldDocument3 pagesAdaptation and Survival: of Organisms in The Changing WorldAyka CantorNo ratings yet
- SOAL PAS KLA X +kunciDocument8 pagesSOAL PAS KLA X +kunciIlyasha Vito DivasNo ratings yet
- Using Product From Canned Tuna Factory Indonesia For Cooking SchoolDocument8 pagesUsing Product From Canned Tuna Factory Indonesia For Cooking SchoolNirvanajunctureNo ratings yet
- Why We Should Tighten Measures OnDocument1 pageWhy We Should Tighten Measures OnFaiz Hagane NatsuNo ratings yet
- Uki and The Ghostburrow (Kieran Larwood David Wyatt)Document218 pagesUki and The Ghostburrow (Kieran Larwood David Wyatt)Ali AbbasNo ratings yet
- Tropical Seaweeds For Human Food, Their Cultivation and Its Effect OnDocument8 pagesTropical Seaweeds For Human Food, Their Cultivation and Its Effect OnJorge Rodríguez VásquezNo ratings yet
- AquaTech A Smart Fishing Farming Automation and Monitoring AppDocument8 pagesAquaTech A Smart Fishing Farming Automation and Monitoring AppIOER International Multidisciplinary Research Journal ( IIMRJ)No ratings yet
- U.S. T R Application For Formal Measurement Services: I. ApplicabilityDocument2 pagesU.S. T R Application For Formal Measurement Services: I. Applicabilitys0606No ratings yet
- Gudang BSDocument5 pagesGudang BSWahyu Nisa AlseraNo ratings yet
- Family ApogonidaeDocument2 pagesFamily ApogonidaelararariingNo ratings yet
- United States Standards For Grades of Frozen Fish Fillet Blocks Scope and Product DescriptionDocument8 pagesUnited States Standards For Grades of Frozen Fish Fillet Blocks Scope and Product DescriptionJackNo ratings yet