You might also like
- Art 06Document4 pagesArt 06Luis Eduardo Zúñiga MaureiraNo ratings yet
- Nettoetal RBP2007Document13 pagesNettoetal RBP2007PatriciaNo ratings yet
- 3 34 Climate Role in Distribution of Cretaceous CrocodyliformesDocument11 pages3 34 Climate Role in Distribution of Cretaceous CrocodyliformesKanekoNo ratings yet
- The Anatomy of A 'Suture Zone' in Amazonian Butter Ies: A Coalescent-Based Test For Vicariant Geographic Divergence and SpeciationDocument20 pagesThe Anatomy of A 'Suture Zone' in Amazonian Butter Ies: A Coalescent-Based Test For Vicariant Geographic Divergence and SpeciationDaniel RodriguezNo ratings yet
- Alcantara Etal 2018 Velloziaceae DiversificationDocument16 pagesAlcantara Etal 2018 Velloziaceae Diversificationdrinkto remenberNo ratings yet
- Unique Cretaceous EcosystemDocument15 pagesUnique Cretaceous EcosystemAlfredo Di Stéfano LaulhéNo ratings yet
- ForamDocument14 pagesForamMeity Irlani AlaiinicanNo ratings yet
- Tafonomía de La Gran Hondonada (Formación Sarmiento, Edad-Mamífero Mustersense, Eoceno Medio) Chubut, ArgentinaDocument16 pagesTafonomía de La Gran Hondonada (Formación Sarmiento, Edad-Mamífero Mustersense, Eoceno Medio) Chubut, ArgentinaAmalia VillafañeNo ratings yet
- 2012 Garcia Massini Et Al - Fungi San AgustinDocument8 pages2012 Garcia Massini Et Al - Fungi San AgustinAlan ChanningNo ratings yet
- Filogeografia PatagoniaDocument20 pagesFilogeografia PatagoniaJuli MartínNo ratings yet
- Menezes Et Al 2017Document16 pagesMenezes Et Al 2017Igor RosárioNo ratings yet
- A History of The Biogeography of Amazonian FishesDocument19 pagesA History of The Biogeography of Amazonian FishesJair ChristianNo ratings yet
- 1 s2.0 S0016699507000101 Main - 3Document13 pages1 s2.0 S0016699507000101 Main - 3Akrem SoltaniNo ratings yet
- RUIZ BUSTOS 1999 - Biostratigraphy of The Continental Deposits in The Granada, Guadix and Baza Basins (Betic Cordillera)Document22 pagesRUIZ BUSTOS 1999 - Biostratigraphy of The Continental Deposits in The Granada, Guadix and Baza Basins (Betic Cordillera)orcenseNo ratings yet
- Revista Mexicana de BiodiversidadDocument19 pagesRevista Mexicana de BiodiversidadAdriana VitoloNo ratings yet
- Lontras Shale (Paraná Basin, Brazil)Document14 pagesLontras Shale (Paraná Basin, Brazil)Crocodilo VasconcelosNo ratings yet
- Equipo 1. Raiza, Lilean, Axel EtcDocument18 pagesEquipo 1. Raiza, Lilean, Axel Etcraizah.rodriguez03No ratings yet
- Arribas HienaDocument19 pagesArribas HienaYsNo ratings yet
- Cardoso 2009Document18 pagesCardoso 2009Liche Puello CaballeroNo ratings yet
- Lizards, Snakes, and Amphisbaenians From The Quaternary Sand Dunes of The Middle Rio Sao Francisco, Bahia, BrazilDocument12 pagesLizards, Snakes, and Amphisbaenians From The Quaternary Sand Dunes of The Middle Rio Sao Francisco, Bahia, Braziluseparadown downNo ratings yet
- Pleistocene Mammals of MexicoDocument52 pagesPleistocene Mammals of MexicoISAAC CAMPOS HERNANDEZNo ratings yet
- Past Climate-Driven Range Shifts Structuring IntraDocument14 pagesPast Climate-Driven Range Shifts Structuring IntraemesethNo ratings yet
- Vicariance - Engl 363 Definition Final Draft - Alisa Hernandez RevisedDocument6 pagesVicariance - Engl 363 Definition Final Draft - Alisa Hernandez Revisedapi-610323824No ratings yet
- Carmona Et Al 2006 (Variability of The Glossifungites Ichnofacies) PDFDocument13 pagesCarmona Et Al 2006 (Variability of The Glossifungites Ichnofacies) PDFSebastian CarvajalinoNo ratings yet
- Yacobaccio, Hugo Et Al. (2016) - Habitats of Ancient Hunter-Gatherers in The Puna. Resilience and Discontinuities During The HoloceneDocument9 pagesYacobaccio, Hugo Et Al. (2016) - Habitats of Ancient Hunter-Gatherers in The Puna. Resilience and Discontinuities During The HoloceneTreder Jauregui CassiaNo ratings yet
- Quaternary International: Hugo D. YacobaccioDocument7 pagesQuaternary International: Hugo D. YacobaccioMartin MarchiNo ratings yet
- Facies Control On Seismites in An Alluvial-Aeolian System The Pliocene Dunefield of The Teruel Half-Graben Basin (Eastern Spain)Document16 pagesFacies Control On Seismites in An Alluvial-Aeolian System The Pliocene Dunefield of The Teruel Half-Graben Basin (Eastern Spain)Johann WolfNo ratings yet
- Bacon 2013Document14 pagesBacon 2013lsapienciaNo ratings yet
- The Stratigraphic and Paleoenvironmental Significance of The Regressive Monte Observación Member, Early Miocene of The Austral Basin, PatagoniaDocument23 pagesThe Stratigraphic and Paleoenvironmental Significance of The Regressive Monte Observación Member, Early Miocene of The Austral Basin, PatagoniaPablo ZuritaNo ratings yet
- Zlab 045Document24 pagesZlab 045Ridwan RahmanNo ratings yet
- Sanmartin2012 EVOO OnlineDocument15 pagesSanmartin2012 EVOO Onlineluis enrique galeana barreraNo ratings yet
- The Importance of The Andes in The Evolutionary RaDocument16 pagesThe Importance of The Andes in The Evolutionary RaAryel MoraisNo ratings yet
- 2010 - Pacheco Etal (Hard Bottom Succession of Subtidal Epibenthic Communities)Document8 pages2010 - Pacheco Etal (Hard Bottom Succession of Subtidal Epibenthic Communities)FrancoNo ratings yet
- Ecology of Syllidae (Annelida: Polychaeta) From Shallow Rocky Environments in The Cantabrian Sea (South Bay of Biscay)Document11 pagesEcology of Syllidae (Annelida: Polychaeta) From Shallow Rocky Environments in The Cantabrian Sea (South Bay of Biscay)garidaki8No ratings yet
- Nat Land 1974Document126 pagesNat Land 1974Juan Guzmán SantosNo ratings yet
- Reimann, C. Et Al. (2006)Document4 pagesReimann, C. Et Al. (2006)williams mataNo ratings yet
- Lagerstätte Effect Drives Notosuchian DiversityDocument11 pagesLagerstätte Effect Drives Notosuchian DiversityCrocodilo VasconcelosNo ratings yet
- 10 1016@j Yqres 2004 02 010Document12 pages10 1016@j Yqres 2004 02 010Laura Sofia Chinchilla MoralesNo ratings yet
- CICCIOLI, Et Al., (2015)Document21 pagesCICCIOLI, Et Al., (2015)kimberly salomeNo ratings yet
- The Evolutionary History of Fouquieriaceae (Ericales) : Biogeography, Growth Habit, Habitat Colonization, and Chromosome EvolutionDocument11 pagesThe Evolutionary History of Fouquieriaceae (Ericales) : Biogeography, Growth Habit, Habitat Colonization, and Chromosome EvolutionMarcela M MNo ratings yet
- Phylogeography of The Chilean Red Cricket Cratomelus Cryptic Diversity in Central ChileDocument16 pagesPhylogeography of The Chilean Red Cricket Cratomelus Cryptic Diversity in Central ChileJoaquín Eduardo Sepúlveda AstudilloNo ratings yet
- Article: Biota Neotropica 21 (1) : E20200985, 2021Document6 pagesArticle: Biota Neotropica 21 (1) : E20200985, 2021Camila MilNo ratings yet
- GIBERT Et Al 1999 - Plio-Pleistocene Deposits of The Orce Region (SE Spain) Geology and AgeDocument18 pagesGIBERT Et Al 1999 - Plio-Pleistocene Deposits of The Orce Region (SE Spain) Geology and AgeorcenseNo ratings yet
- Aldeias Et Al-2016-Geoarchaeology PDFDocument20 pagesAldeias Et Al-2016-Geoarchaeology PDFJorge Alonso Rodriguez MoralesNo ratings yet
- Fishing Strategies Among Prehistoric Populations at Saquarema Lagoonal Complex, Rio de Janeiro, BrazilDocument15 pagesFishing Strategies Among Prehistoric Populations at Saquarema Lagoonal Complex, Rio de Janeiro, Brazilmarcia guimaraesNo ratings yet
- Understanding The Evolutionary History of A High Andean Endemic The Ecuadorian Hillstar Oreotrochilus ChimborazoDocument15 pagesUnderstanding The Evolutionary History of A High Andean Endemic The Ecuadorian Hillstar Oreotrochilus ChimborazoDavid AndradeNo ratings yet
- Bioregiones Andes TropicalesDocument6 pagesBioregiones Andes TropicalesLilia SierraNo ratings yet
- Daniels Etal 2019 Revised Chronostratigraphic Cretaceous Magallanes Ultima EsperanzaDocument16 pagesDaniels Etal 2019 Revised Chronostratigraphic Cretaceous Magallanes Ultima EsperanzaSebastian Kaempfe DroguettNo ratings yet
- The Trace Fossil MesozoicCenozoic PDFDocument497 pagesThe Trace Fossil MesozoicCenozoic PDFCamila Vega100% (2)
- UntitledDocument19 pagesUntitledAstolfo Gomes de Mello AraujoNo ratings yet
- Brachiopod and Bivalve Ecology in The Late Triassi PDFDocument32 pagesBrachiopod and Bivalve Ecology in The Late Triassi PDFasmatiNo ratings yet
- Paleobiogeography: Presented by - Jogya Jyoti Medok Jayshree Bora Abdul Waris Rohit Rajak Nilotpal ShyamDocument28 pagesPaleobiogeography: Presented by - Jogya Jyoti Medok Jayshree Bora Abdul Waris Rohit Rajak Nilotpal ShyamAnurag pritamNo ratings yet
- 1 s2.0 S0169555X23002842 MainDocument65 pages1 s2.0 S0169555X23002842 MainCarlos Saez MartinezNo ratings yet
- Bats and Elevation 2010Document14 pagesBats and Elevation 2010Johnny BravoNo ratings yet
- Geo2015 11 14Document6 pagesGeo2015 11 14HalsandNo ratings yet
- Paleoenvironment and Age of Los Monos Formation (Devonian), Tarija Basin, Argentina and BoliviaDocument36 pagesPaleoenvironment and Age of Los Monos Formation (Devonian), Tarija Basin, Argentina and BoliviaCristian Mauricio Calle LinconNo ratings yet
- 0research Proposal SAJAD - Edited.editedDocument4 pages0research Proposal SAJAD - Edited.editedMir Sajjad HüSsàïñNo ratings yet
- Paleontological Constraints On Successive Paleogeographic Positions of Precordillera Terrane During The Early PaleozoicDocument23 pagesPaleontological Constraints On Successive Paleogeographic Positions of Precordillera Terrane During The Early PaleozoicVxx XxNo ratings yet
- Bones, Clones, and Biomes: The History and Geography of Recent Neotropical MammalsFrom EverandBones, Clones, and Biomes: The History and Geography of Recent Neotropical MammalsNo ratings yet
- Ecological Informatics: B. Zaragozí, A. Belda, P. Giménez, J.T. Navarro, A. BonetDocument6 pagesEcological Informatics: B. Zaragozí, A. Belda, P. Giménez, J.T. Navarro, A. BonetSebas CirignoliNo ratings yet
- Zoo Biology, 17, 333-341.Document9 pagesZoo Biology, 17, 333-341.Sebas CirignoliNo ratings yet
- Unpacking Equity For Protected Area Conservation: Kate Schreckenberg, Phil Franks, Adrian Martin and Barbara LangDocument16 pagesUnpacking Equity For Protected Area Conservation: Kate Schreckenberg, Phil Franks, Adrian Martin and Barbara LangSebas CirignoliNo ratings yet
- Herpailurus Yagouaroundi,: Assessment By: Caso, A., de Oliveira, T. & Carvajal, S.VDocument12 pagesHerpailurus Yagouaroundi,: Assessment By: Caso, A., de Oliveira, T. & Carvajal, S.VSebas CirignoliNo ratings yet
- Zoo Biology, 17, 369-371.Document3 pagesZoo Biology, 17, 369-371.Sebas CirignoliNo ratings yet
- Zoo Biology, 17, 231-244.Document14 pagesZoo Biology, 17, 231-244.Sebas CirignoliNo ratings yet
- Zoo Biology, 22, 489-496.Document8 pagesZoo Biology, 22, 489-496.Sebas CirignoliNo ratings yet
- Zoo Biology, 17, 373-377.Document5 pagesZoo Biology, 17, 373-377.Sebas CirignoliNo ratings yet
- Zoo Biology, 16, 549-550.Document2 pagesZoo Biology, 16, 549-550.Sebas CirignoliNo ratings yet
- Coming in Out of The Cold: Animal Keeping: in Temperate Zone ZoosDocument3 pagesComing in Out of The Cold: Animal Keeping: in Temperate Zone ZoosSebas CirignoliNo ratings yet
- Zoo Biology, 17, 167-180.Document14 pagesZoo Biology, 17, 167-180.Sebas CirignoliNo ratings yet
- Zoo Biology, 17, 167-180.Document14 pagesZoo Biology, 17, 167-180.Sebas CirignoliNo ratings yet
- Zoo Biology, 16, 549-550.Document2 pagesZoo Biology, 16, 549-550.Sebas CirignoliNo ratings yet
- Zoo Biology, 16, 377-389.Document13 pagesZoo Biology, 16, 377-389.Sebas CirignoliNo ratings yet
- Zoo Biology, 16, 103-106.Document4 pagesZoo Biology, 16, 103-106.Sebas CirignoliNo ratings yet
- Zoo Biology, 16, 103-106.Document4 pagesZoo Biology, 16, 103-106.Sebas CirignoliNo ratings yet
- Larson 1997 - Taxonomic Re-Evaluation of The JaguarDocument14 pagesLarson 1997 - Taxonomic Re-Evaluation of The JaguarannesmartinsNo ratings yet
- Zoo Biology, 16, 377-389.Document13 pagesZoo Biology, 16, 377-389.Sebas CirignoliNo ratings yet
- Zoo Biology, 16, 301-307.Document7 pagesZoo Biology, 16, 301-307.Sebas CirignoliNo ratings yet
- Larson 1997 - Taxonomic Re-Evaluation of The JaguarDocument14 pagesLarson 1997 - Taxonomic Re-Evaluation of The JaguarannesmartinsNo ratings yet
- Zoo Biology, 15, 509-524.Document16 pagesZoo Biology, 15, 509-524.Sebas CirignoliNo ratings yet
- Zoo Biology, 16, 71-93.Document13 pagesZoo Biology, 16, 71-93.Sebas CirignoliNo ratings yet
- Zoo Biology, 16, 1-2.Document2 pagesZoo Biology, 16, 1-2.Sebas CirignoliNo ratings yet
- Zoo Biology, 15, 491-497.Document7 pagesZoo Biology, 15, 491-497.Sebas CirignoliNo ratings yet
- Western North American Naturalist, 61 (4), 501-502.Document2 pagesWestern North American Naturalist, 61 (4), 501-502.Sebas CirignoliNo ratings yet
- Zoo Biology, 15, 525-537.Document13 pagesZoo Biology, 15, 525-537.Sebas CirignoliNo ratings yet
- Web Ecology, 3, 56-69.Document14 pagesWeb Ecology, 3, 56-69.Sebas CirignoliNo ratings yet
- Zoo Biology, 15, 481-490.Document11 pagesZoo Biology, 15, 481-490.Sebas CirignoliNo ratings yet
- Acta Parasitologica, 49 (4)Document11 pagesActa Parasitologica, 49 (4)Sebas CirignoliNo ratings yet
- Avian Diseases, 46, 754-756 - Rhea AmericanaDocument3 pagesAvian Diseases, 46, 754-756 - Rhea AmericanaSebas CirignoliNo ratings yet
- Projet Report Flash Flood of UttarakhandDocument20 pagesProjet Report Flash Flood of UttarakhandDiyasha GangulyNo ratings yet
- TN FC PDFDocument2 pagesTN FC PDFRamesh NatarajanNo ratings yet
- Summative Test 1&2Document3 pagesSummative Test 1&2Takeshi KuniiNo ratings yet
- ICSE Solutions For Class 9 Geography - Insolation +Document20 pagesICSE Solutions For Class 9 Geography - Insolation +Home MadeNo ratings yet
- DRRM The Albay Experience and Sustainable PlanDocument65 pagesDRRM The Albay Experience and Sustainable PlanZeno Silva100% (1)
- Dry and Wet Spell Probability Analysis by Markov Chain Model at Parbhani, MaharashtraDocument5 pagesDry and Wet Spell Probability Analysis by Markov Chain Model at Parbhani, MaharashtraUnnati SonawaneNo ratings yet
- Learning Area Grade Level Quarter Date I. Lesson TitleDocument22 pagesLearning Area Grade Level Quarter Date I. Lesson TitleEnohoj YamNo ratings yet
- NCERT Solutions For Class 9: Chapter 1 India - Size and LocationDocument7 pagesNCERT Solutions For Class 9: Chapter 1 India - Size and LocationArdhendu SamantaNo ratings yet
- Sambhar LakeDocument3 pagesSambhar LakeAditya BhandariNo ratings yet
- ArtigoDocument18 pagesArtigoMariane de LizNo ratings yet
- I.Objectives A. Content StandardDocument3 pagesI.Objectives A. Content StandardJe-Ann Descalsota RelotaNo ratings yet
- Stages For Construction W3b PDFDocument8 pagesStages For Construction W3b PDFfarrahfarinaNo ratings yet
- The Malaysian RainforestDocument3 pagesThe Malaysian RainforestVednivead100% (2)
- Science 7 Q4 M1 Day 1Document33 pagesScience 7 Q4 M1 Day 1Shelvie Dy IcoNo ratings yet
- Hurricanes and TyphoonsDocument33 pagesHurricanes and TyphoonsDella Pantaleon ParasNo ratings yet
- GEO 387 W4 Thermodynamics Dry MyDocument38 pagesGEO 387 W4 Thermodynamics Dry MyLmaoNo ratings yet
- APC - OCE551 - Air Pollution and Control Engineering - UnitII-MeteorologyDocument14 pagesAPC - OCE551 - Air Pollution and Control Engineering - UnitII-Meteorologyaruna mahadevan67% (9)
- Indian Physical GeographyDocument108 pagesIndian Physical GeographyMithunNo ratings yet
- Prologue To Jurassic Park by Michael CrichtonDocument1 pagePrologue To Jurassic Park by Michael CrichtonBurt Gummer100% (7)
- Lesson 1. What Is An EcosystemDocument10 pagesLesson 1. What Is An EcosystemLabib Kamal Year 11No ratings yet
- Typhoon KetsanaDocument3 pagesTyphoon Ketsanadan teNo ratings yet
- Landslide and Sinkhole 2Document27 pagesLandslide and Sinkhole 2merrylace sanjuanNo ratings yet
- Bab 1Document5 pagesBab 1AjengNo ratings yet
- Niwrmp Draft ReportDocument90 pagesNiwrmp Draft ReportMogesNo ratings yet
- LG Chem Design - Guideline - ExternalDocument2 pagesLG Chem Design - Guideline - ExternalAlshuqaiq LaboratoryNo ratings yet
- Forests of KarnatakaDocument10 pagesForests of Karnatakasanjay kumarNo ratings yet
- Climate Responsive ArchitectureDocument27 pagesClimate Responsive ArchitectureRavnish BatthNo ratings yet
- Conner 1957Document4 pagesConner 1957Rafael E. Becemberg LNo ratings yet
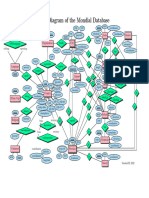
- Mondial ERDocument1 pageMondial ERPLK 225No ratings yet
- Saturated Greenhouse Effect TheoryDocument5 pagesSaturated Greenhouse Effect TheoryDiannaCC100% (1)