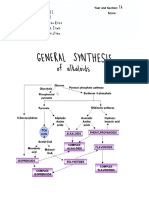
You might also like
- Recent Advances in Polyphenol ResearchFrom EverandRecent Advances in Polyphenol ResearchHeidi HalbwirthNo ratings yet
- Biotechnology of Terpenoid Production from Microbial Cell FactoriesFrom EverandBiotechnology of Terpenoid Production from Microbial Cell FactoriesNo ratings yet
- Labdanes of Natural Origin-Biological ActivitiesDocument24 pagesLabdanes of Natural Origin-Biological Activitiestheyuri@tlen.plNo ratings yet
- Terpenoids PaperDocument24 pagesTerpenoids PaperKingsley UgwuNo ratings yet
- Natural Products ChemistryDocument80 pagesNatural Products ChemistrychimnecheremukachukwuNo ratings yet
- TerpenoidsDocument146 pagesTerpenoidsAnonymous TCbZigVqNo ratings yet
- Biosynthesis of Plant Secondary MetabolitesDocument34 pagesBiosynthesis of Plant Secondary MetabolitesPuvaneswary LoganathanNo ratings yet
- 8 - TerpenoidDocument95 pages8 - TerpenoidTim IrmaNo ratings yet
- TerpeneDocument11 pagesTerpeneweweweweNo ratings yet
- Use of Terpenoids As Natural FlavouringDocument8 pagesUse of Terpenoids As Natural FlavouringLuis GracianoNo ratings yet
- Plant Secondary MetabolismDocument108 pagesPlant Secondary MetabolismanamambriNo ratings yet
- 6 PhenylpropanoidsDocument15 pages6 Phenylpropanoidskusum AkkiNo ratings yet
- Group 03 (2A) 2Document8 pagesGroup 03 (2A) 2-No ratings yet
- Natural Products: Dr. Tukiran, M.SiDocument18 pagesNatural Products: Dr. Tukiran, M.SiSangeetha PrakashNo ratings yet
- SCL Organik 4 Group 1Document13 pagesSCL Organik 4 Group 1Wan HafizaNo ratings yet
- Trabalho ChesyDocument14 pagesTrabalho ChesyMario AraujoNo ratings yet
- 8 - Secondary Metabolites - TERPENOIDS ALKALOIDS PHENOLICS PDFDocument82 pages8 - Secondary Metabolites - TERPENOIDS ALKALOIDS PHENOLICS PDFFCiênciasNo ratings yet
- Unit 10Document24 pagesUnit 10debksaha2050No ratings yet
- Antioxidan 1Document43 pagesAntioxidan 1pardeepbthNo ratings yet
- Secondary Metabolites B.sc. Botany 3rd by Dr. Meenu Gupta 2Document18 pagesSecondary Metabolites B.sc. Botany 3rd by Dr. Meenu Gupta 2svaidyaNo ratings yet
- Made by ..Naila Gul MSC Previous R.No. 23Document23 pagesMade by ..Naila Gul MSC Previous R.No. 23Asif Ali100% (2)
- Organic Chemistry Experiment Report Phytochemical Test On The Extract of Wild Ginger (Curcuma Zanthorrhiza)Document18 pagesOrganic Chemistry Experiment Report Phytochemical Test On The Extract of Wild Ginger (Curcuma Zanthorrhiza)auraimanaNo ratings yet
- Introduction of TerpenoidsDocument17 pagesIntroduction of TerpenoidsAbdul WahhabNo ratings yet
- Pharmacognosy AssignmentDocument22 pagesPharmacognosy AssignmentJayed SadnanNo ratings yet
- Ncomms 1327Document9 pagesNcomms 1327Chandra ShekharNo ratings yet
- Plant Secondary MetabolitesDocument36 pagesPlant Secondary MetabolitesvishnuNo ratings yet
- Terp e NoidsDocument23 pagesTerp e Noidsrusbianto wijayaNo ratings yet
- Phytochemical Investigation On The Ethanol Extract of The Aerial Parts of Laggera TomentosaDocument15 pagesPhytochemical Investigation On The Ethanol Extract of The Aerial Parts of Laggera Tomentosainternational journal of modern chemistry and applied scienceNo ratings yet
- Effects of ABA On Primary Terpenoids and THC in Cannabis Sativa at Flowering StageDocument9 pagesEffects of ABA On Primary Terpenoids and THC in Cannabis Sativa at Flowering StageJaviercm10No ratings yet
- Terpenes and TerpenoidsDocument10 pagesTerpenes and TerpenoidsMarwah100% (1)
- Biotechnological Process of Isoperno by Extraction From Pine PatulaDocument13 pagesBiotechnological Process of Isoperno by Extraction From Pine Patulajuan labradorNo ratings yet
- Fermentation: Metabolic BiodiversityDocument10 pagesFermentation: Metabolic BiodiversityEduar Moreno LondoñoNo ratings yet
- Approximately 1500 Colored CompoundsDocument15 pagesApproximately 1500 Colored CompoundsAnur SinglaNo ratings yet
- Neil C. Bruce - AlkaloidsDocument11 pagesNeil C. Bruce - AlkaloidsHumiopNo ratings yet
- OC-V p5Document14 pagesOC-V p5Derek Ross0% (1)
- Terpenes and SteroidsDocument38 pagesTerpenes and SteroidsZabrina Tan CuaNo ratings yet
- Phytochemistry of The Plant (Senna Tora Linn)Document22 pagesPhytochemistry of The Plant (Senna Tora Linn)Efeturi Ovie KennedyNo ratings yet
- MenThol ReviewDocument13 pagesMenThol ReviewLuke RossNo ratings yet
- Bio Synthesis of Organic CompoundsDocument18 pagesBio Synthesis of Organic Compoundshgfhfg3621No ratings yet
- Isoquinoline - Lec6 PDFDocument16 pagesIsoquinoline - Lec6 PDFHassan mohamad Al-bayateNo ratings yet
- AlkaloidsDocument7 pagesAlkaloidsLizbeth Aura CebrianNo ratings yet
- UntitledDocument69 pagesUntitledCoyme Rey JohnNo ratings yet
- Biosintesis of TerpenoidDocument19 pagesBiosintesis of TerpenoidIntanFrinaUtamiyantiNo ratings yet
- Secondary Metabolites Pankaj PresentationDocument97 pagesSecondary Metabolites Pankaj PresentationPankaj Pant100% (1)
- Phytochemistry of AlkaloidsDocument39 pagesPhytochemistry of AlkaloidsThamer JaberNo ratings yet
- Phenylpropanoids1Document43 pagesPhenylpropanoids1AdzimahNo ratings yet
- Org - Unit 4Document29 pagesOrg - Unit 4lijyohannesmekonnen7No ratings yet
- Biotechnology For The Production of Essential Oils, Avours and Volatile Isolates. A ReviewDocument20 pagesBiotechnology For The Production of Essential Oils, Avours and Volatile Isolates. A ReviewrobertetNo ratings yet
- Medicines 03 00025Document16 pagesMedicines 03 00025Syed Iftekhar AlamNo ratings yet
- Bergman and Phillips 2020 Phytochemistry Reviews OnlineDocument27 pagesBergman and Phillips 2020 Phytochemistry Reviews OnlineLeen OdehNo ratings yet
- Algunas Reacciones de EugenolDocument31 pagesAlgunas Reacciones de Eugenoldan martinezNo ratings yet
- Phyto AlexinsDocument11 pagesPhyto AlexinsanastatataNo ratings yet
- Derivatives of AlcoholDocument5 pagesDerivatives of AlcoholEcho MoralesNo ratings yet
- Pendahuluan, Metabolit Primer Dan Sekunder PDFDocument85 pagesPendahuluan, Metabolit Primer Dan Sekunder PDFViena CynthiaNo ratings yet
- The Chemistry of Natural Product Plant Secondary MetabolitesDocument9 pagesThe Chemistry of Natural Product Plant Secondary MetabolitesImdad HussainNo ratings yet
- 05-Terpenos y EsteroidesDocument23 pages05-Terpenos y EsteroidesAle VillaNo ratings yet
- Alcaloides A 2023Document76 pagesAlcaloides A 2023Alex osorioNo ratings yet
- PhenolDocument4 pagesPhenolAbeer AbdelganeNo ratings yet
- Terpenoids From EoDocument22 pagesTerpenoids From EoIsidora NikolicNo ratings yet
- AlkaloidsDocument9 pagesAlkaloidsHari sankar MuniNo ratings yet
- Ephrins: Ru Diger KleinDocument6 pagesEphrins: Ru Diger Kleinmanoj_rkl_07No ratings yet
- 2,4-Thiazolidinedione As Antimicrobial and Cytotoxic AgentsDocument10 pages2,4-Thiazolidinedione As Antimicrobial and Cytotoxic Agentsmanoj_rkl_07No ratings yet
- Evolution of Ecosystems - Terrestrial PDFDocument4 pagesEvolution of Ecosystems - Terrestrial PDFmanoj_rkl_07No ratings yet
- Evolution of Genome Organizn1699-001-000 PDFDocument5 pagesEvolution of Genome Organizn1699-001-000 PDFmanoj_rkl_07No ratings yet
- Evolution of Development A0001661-001-000 PDFDocument4 pagesEvolution of Development A0001661-001-000 PDFmanoj_rkl_07No ratings yet
- Ca Binding Proteins A0001347-001-000 PDFDocument8 pagesCa Binding Proteins A0001347-001-000 PDFmanoj_rkl_07No ratings yet
- DNA Damage: Paul W DoetschDocument7 pagesDNA Damage: Paul W Doetschmanoj_rkl_07No ratings yet
- Electron Carriers PDFDocument8 pagesElectron Carriers PDFmanoj_rkl_07100% (1)
- BMP Antags &nural Inducna0000805-001-000 PDFDocument5 pagesBMP Antags &nural Inducna0000805-001-000 PDFmanoj_rkl_07No ratings yet
- Cyanogenesis in Higher Plant and InsectsDocument3 pagesCyanogenesis in Higher Plant and Insectsmanoj_rkl_07No ratings yet
- Plant Macro-And Micronutrient MineralsDocument5 pagesPlant Macro-And Micronutrient Mineralsmanoj_rkl_07No ratings yet
- Capillary Blotting of RNA and DNA Gels PDFDocument3 pagesCapillary Blotting of RNA and DNA Gels PDFmanoj_rkl_07No ratings yet
- Electroporation: Jac A NickoloffDocument3 pagesElectroporation: Jac A Nickoloffmanoj_rkl_07No ratings yet
- DNA Repair by Reversal of Damage PDFDocument8 pagesDNA Repair by Reversal of Damage PDFmanoj_rkl_07100% (1)
- Heavy Metal Adaptation PDFDocument4 pagesHeavy Metal Adaptation PDFmanoj_rkl_07No ratings yet
- Forward-Cristopher Reeve PDFDocument1 pageForward-Cristopher Reeve PDFmanoj_rkl_07No ratings yet
- Calibration of A Transillumanator For UV Fixation of DNA To PDFDocument2 pagesCalibration of A Transillumanator For UV Fixation of DNA To PDFmanoj_rkl_07No ratings yet
- Genetic Code Introduction PDFDocument10 pagesGenetic Code Introduction PDFmanoj_rkl_07No ratings yet
- Phyllosphere PDFDocument8 pagesPhyllosphere PDFmanoj_rkl_07No ratings yet
- FGF1Document9 pagesFGF1manoj_rkl_07No ratings yet
- Dideoxy Sequencing of DNA PDFDocument16 pagesDideoxy Sequencing of DNA PDFmanoj_rkl_07No ratings yet
- Closteroviridae: Historical PerspectiveDocument6 pagesClosteroviridae: Historical Perspectivemanoj_rkl_07No ratings yet
- Immunologival Tolerance Therpeutic Induction PDFDocument6 pagesImmunologival Tolerance Therpeutic Induction PDFmanoj_rkl_07No ratings yet
- Flocyt Anal Nstem Cel in DevDocument11 pagesFlocyt Anal Nstem Cel in Devmanoj_rkl_07No ratings yet
- Calibration of A Transillumanator For UV Fixation of DNA To PDFDocument2 pagesCalibration of A Transillumanator For UV Fixation of DNA To PDFmanoj_rkl_07No ratings yet
- Heavy Metal Adaptation PDFDocument4 pagesHeavy Metal Adaptation PDFmanoj_rkl_07No ratings yet
- Gertrude Belle Elion PDFDocument1 pageGertrude Belle Elion PDFmanoj_rkl_07No ratings yet
- Root Nodules (Rhizobium Legumes) PDFDocument2 pagesRoot Nodules (Rhizobium Legumes) PDFmanoj_rkl_07No ratings yet
- Dideoxy Sequencing of DNA PDFDocument16 pagesDideoxy Sequencing of DNA PDFmanoj_rkl_07No ratings yet
- Personal ComputerDocument3 pagesPersonal ComputerDan Mark IsidroNo ratings yet
- Fix LHA Whole PagesDocument81 pagesFix LHA Whole PagesvuonghhNo ratings yet
- Gulf Case Study SolnDocument9 pagesGulf Case Study SolnHarsh SNo ratings yet
- Flyweis Services Pvt. LTDDocument11 pagesFlyweis Services Pvt. LTDFlyweis TechnologyNo ratings yet
- PhysioEx Exercise 1 Activity 1Document3 pagesPhysioEx Exercise 1 Activity 1edvin merida proNo ratings yet
- Check Out The Buyers Guide On FacebookDocument28 pagesCheck Out The Buyers Guide On FacebookCoolerAdsNo ratings yet
- User Manual - Wellwash ACDocument99 pagesUser Manual - Wellwash ACAlexandrNo ratings yet
- Injection Nozzle Opening Pressure: Lmbo QFKD Fkpqor'qflkDocument1 pageInjection Nozzle Opening Pressure: Lmbo QFKD Fkpqor'qflkAlexanderNo ratings yet
- Python PyDocument19 pagesPython Pyakhilesh kr bhagatNo ratings yet
- Macro Economics A2 Level Notes Book PDFDocument33 pagesMacro Economics A2 Level Notes Book PDFMustafa Bilal50% (2)
- UGC NET Paper I PreviousDocument16 pagesUGC NET Paper I PreviousKirran Khumar GollaNo ratings yet
- W1 - V1 MultipleWorksheets SolnDocument3 pagesW1 - V1 MultipleWorksheets SolnAKHIL RAJ SNo ratings yet
- Genstat Release 10.3de (Pc/Windows 7) 28 May 2012 06:35:59Document6 pagesGenstat Release 10.3de (Pc/Windows 7) 28 May 2012 06:35:59Anna Nur HidayatiNo ratings yet
- Fashion Goes VirtualDocument1 pageFashion Goes VirtualJessica MichaultNo ratings yet
- BTL - 5000 SWT - Service Manual PDFDocument158 pagesBTL - 5000 SWT - Service Manual PDFNuno Freitas BastosNo ratings yet
- Syllabus Math 305Document4 pagesSyllabus Math 305Nataly SusanaNo ratings yet
- Anzsco SearchDocument6 pagesAnzsco SearchytytNo ratings yet
- Nanomedicine Lecture 2007Document59 pagesNanomedicine Lecture 200778912071No ratings yet
- CFA L1 Ethics Questions and AnswersDocument94 pagesCFA L1 Ethics Questions and AnswersMaulik PatelNo ratings yet
- Consumer Price SummaryDocument5 pagesConsumer Price SummaryKJ HiramotoNo ratings yet
- Opening The Third EyeDocument13 pagesOpening The Third EyekakamacgregorNo ratings yet
- Engineering ChemistryDocument21 pagesEngineering Chemistryjai kumarNo ratings yet
- EverServ 7700 M77XX Quick Reference GuideDocument2 pagesEverServ 7700 M77XX Quick Reference GuidetangocharliepdxNo ratings yet
- Esp 1904 A - 70 TPH o & M ManualDocument50 pagesEsp 1904 A - 70 TPH o & M Manualpulakjaiswal85No ratings yet
- 0apageo Catalogue Uk 2022Document144 pages0apageo Catalogue Uk 2022Kouassi JaurèsNo ratings yet
- Reflection Frog 1Document3 pagesReflection Frog 1mariamNo ratings yet
- Pinto pm5 Tif 02Document24 pagesPinto pm5 Tif 02Salem BawazirNo ratings yet
- Chemistry For PhotographersDocument184 pagesChemistry For PhotographersBahar ShoghiNo ratings yet
- Shsa1105 - Unit-III Course MaterialsDocument58 pagesShsa1105 - Unit-III Course Materialssivanikesh bonagiriNo ratings yet
- Analysis of Mozarts k.475Document2 pagesAnalysis of Mozarts k.475ASPASIA FRAGKOUNo ratings yet