You might also like
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5795)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1091)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (895)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (588)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (345)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (121)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (400)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2259)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (74)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- Hydraulics A320Document101 pagesHydraulics A320KamalVirk100% (6)
- FLUSARC 36kV - 630A - 25ka Maintenance Manual PDFDocument18 pagesFLUSARC 36kV - 630A - 25ka Maintenance Manual PDFJoey Real CabalidaNo ratings yet
- Review of Xanthan Gum Production From Unmodified Starches-2006Document11 pagesReview of Xanthan Gum Production From Unmodified Starches-2006ashuna249396No ratings yet
- ZMQ 9 H 2 CM5 I0 CDocument666 pagesZMQ 9 H 2 CM5 I0 Cashuna249396No ratings yet
- BT 0306Document3 pagesBT 0306ashuna249396No ratings yet
- Xanthan Gum ScifinderDocument15 pagesXanthan Gum Scifinderashuna249396No ratings yet
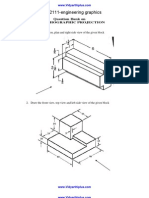
- GE2111-engineering Graphics: 1. Draw The Elevation, Plan and Right Side View of The Given BlockDocument4 pagesGE2111-engineering Graphics: 1. Draw The Elevation, Plan and Right Side View of The Given Blockashuna249396No ratings yet
- Production of Poly (3-Hydroxybutyrate) and Poly (3-Hydroxybutyrate-Co-4-Hydroxybutyrate) by Ralstonia Eutropha From Soybean OilDocument6 pagesProduction of Poly (3-Hydroxybutyrate) and Poly (3-Hydroxybutyrate-Co-4-Hydroxybutyrate) by Ralstonia Eutropha From Soybean Oilashuna249396No ratings yet
- Model-Based Fed-Batch Cultivation of R. Eutropha For Enhanced Biopolymer ProductionDocument8 pagesModel-Based Fed-Batch Cultivation of R. Eutropha For Enhanced Biopolymer Productionashuna249396No ratings yet
- Microbial Production of Polyhydroxybutyrate, A Biodegradable Plastic Using Agro-Industrial Waste ProductsDocument11 pagesMicrobial Production of Polyhydroxybutyrate, A Biodegradable Plastic Using Agro-Industrial Waste Productsashuna249396No ratings yet
- Appl. Environ. Microbiol. 2009 Ibrahim 6222 31Document10 pagesAppl. Environ. Microbiol. 2009 Ibrahim 6222 31ashuna249396No ratings yet
- Important MCQ Colloction For PartDocument154 pagesImportant MCQ Colloction For PartNazia Ali86% (7)
- Acute Management of Burn and Electrical TraumaDocument35 pagesAcute Management of Burn and Electrical TraumaRosa GutiérrezNo ratings yet
- The Neurovascular Bundle of The Anterior Compartment ofDocument30 pagesThe Neurovascular Bundle of The Anterior Compartment ofAnnapurna Bose100% (2)
- Solid Waste Management in SchoolsDocument32 pagesSolid Waste Management in SchoolsRusty TorioNo ratings yet
- SafetyMAP Measuring ManagementDocument4 pagesSafetyMAP Measuring Managementindri anggrainiNo ratings yet
- The Four Common Types of Parenting StylesDocument11 pagesThe Four Common Types of Parenting StylesIka_Dyah_Purwa_1972100% (3)
- Modelling The Transport of Crude Oil in Sandy Soil: Ejikeme Ugwoha, Victor Emeka Amah, Precious Ehis Agharese-AduDocument12 pagesModelling The Transport of Crude Oil in Sandy Soil: Ejikeme Ugwoha, Victor Emeka Amah, Precious Ehis Agharese-AdusfNo ratings yet
- Sewage Disposal System Treatment and Recycling - Group 1Document17 pagesSewage Disposal System Treatment and Recycling - Group 1Rosevie Mae EnriqueNo ratings yet
- History Checklist - Sara AlJasserDocument3 pagesHistory Checklist - Sara AlJassersalman khanNo ratings yet
- PMBOKDocument11 pagesPMBOKshafoooNo ratings yet
- 5.1 Study GuideDocument5 pages5.1 Study GuideChris GayleNo ratings yet
- Same Engines Spare PartsDocument98 pagesSame Engines Spare PartsAdrian Macaya100% (1)
- Slaking SQ Physical Indicator SheetDocument2 pagesSlaking SQ Physical Indicator Sheetqwerty12348No ratings yet
- AS 2122 HK1 DecuongHKI ANH7Document19 pagesAS 2122 HK1 DecuongHKI ANH7Nguyễn Lê KhánhNo ratings yet
- Whsms ManualDocument22 pagesWhsms ManualSayed AbbasNo ratings yet
- MotherDocument11 pagesMotherMuhammad Lucky AlfaridzyNo ratings yet
- Safety Precautions For Handling ChemicalsDocument68 pagesSafety Precautions For Handling ChemicalsRaul FenrandezNo ratings yet
- Sentinel Node Biopsy For CA BreastDocument7 pagesSentinel Node Biopsy For CA Breastwasima1956No ratings yet
- Blok-Aid Sewer Overflow Monitoring and Alarm SystemDocument2 pagesBlok-Aid Sewer Overflow Monitoring and Alarm SystemTomas LarsenNo ratings yet
- 5000L Series CatalogueDocument24 pages5000L Series CataloguekahlaouiNo ratings yet
- Chapter 3: Curriculum Design: The Teacher and The School Curriculum A Guide To Curriculum DevelopmentDocument14 pagesChapter 3: Curriculum Design: The Teacher and The School Curriculum A Guide To Curriculum Developmenthannah mari100% (1)
- 68BDocument5 pages68BJamie Schultz100% (1)
- Lbe Diskusi 16 Tambahan FixDocument4 pagesLbe Diskusi 16 Tambahan FixRayhan Al FaiqNo ratings yet
- Pile Hammer Delmag d12-42Document1 pagePile Hammer Delmag d12-42Jorge Rosero QuevedoNo ratings yet
- Project Planning Template - Castillo JiselleDocument7 pagesProject Planning Template - Castillo Jiselleapi-692396370No ratings yet
- Understanding Building Construction Types - Firefighter Nation PDFDocument3 pagesUnderstanding Building Construction Types - Firefighter Nation PDFRakeshNo ratings yet
- Sikafloor - 325: 2-Part PUR Resin Self Smoothing Screed and Seal CoatDocument6 pagesSikafloor - 325: 2-Part PUR Resin Self Smoothing Screed and Seal Coatthe pilotNo ratings yet
- Clinical Review: Independent Lung Ventilation in Critical CareDocument7 pagesClinical Review: Independent Lung Ventilation in Critical CareIrene Cristina100% (1)