You might also like
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5784)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (890)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (399)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (587)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (265)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (72)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (344)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2219)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (119)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- The Posterior Fossa Cisterns: Albert L. Rhoton, JR., M.DDocument12 pagesThe Posterior Fossa Cisterns: Albert L. Rhoton, JR., M.DIndra HadianditeNo ratings yet
- Classification of dental anomalies and deformitiesDocument15 pagesClassification of dental anomalies and deformitiesAdelina MijaicheNo ratings yet
- Merged Ortho MCQDocument51 pagesMerged Ortho MCQblessy winsletNo ratings yet
- The Vascular SystemDocument7 pagesThe Vascular SystemMiguel GumatayNo ratings yet
- Root Canal Cover UpDocument248 pagesRoot Canal Cover UpVictor Mendes97% (36)
- Zollinger-Ellison Syndrome (Gastrinoma) - Clinical Manifestations and Diagnosis - UpToDateDocument14 pagesZollinger-Ellison Syndrome (Gastrinoma) - Clinical Manifestations and Diagnosis - UpToDateFeer Villarreal100% (1)
- Muscle TissueDocument20 pagesMuscle TissueBrian MirasolNo ratings yet
- Fang and Fur Blood and Bone PDFDocument2 pagesFang and Fur Blood and Bone PDFLuis0% (2)
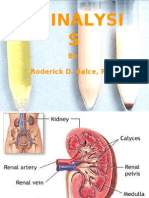
- CLINICAL MICROSCOPY (Urinalysis)Document131 pagesCLINICAL MICROSCOPY (Urinalysis)Roderick95% (19)
- Circulation. Lec On HemodynamicsDocument30 pagesCirculation. Lec On HemodynamicsMudassar RoomiNo ratings yet
- 3 +endocrine+systemDocument41 pages3 +endocrine+systemDew JirawatNo ratings yet
- Oral Hygiene Nursing NoteDocument2 pagesOral Hygiene Nursing NoteLinda AdelinaNo ratings yet
- Choosing A Pre-Adjusted Orthodontic Appliance Prescription For Anterior TeethDocument6 pagesChoosing A Pre-Adjusted Orthodontic Appliance Prescription For Anterior TeethYorisbeth SanNo ratings yet
- The Nervous System: Test Yourself 13.1 (Page 258)Document2 pagesThe Nervous System: Test Yourself 13.1 (Page 258)leeNo ratings yet
- A&PII Test 1Document14 pagesA&PII Test 1Alyssa BattcockNo ratings yet
- LUNG-OVERVIEW (Autosaved)Document132 pagesLUNG-OVERVIEW (Autosaved)Thivashinie Kandy Nazan VelloNo ratings yet
- References: Points Weight Lecture Actual HoursDocument8 pagesReferences: Points Weight Lecture Actual HoursAlyssa SanchezNo ratings yet
- Student Exploration: Circulatory SystemDocument4 pagesStudent Exploration: Circulatory Systemna shadolstalkerNo ratings yet
- Adrenal Gland: Bardelosa, Jesse Gale M. Barretto, Alyssa Nicole 3MT1Document88 pagesAdrenal Gland: Bardelosa, Jesse Gale M. Barretto, Alyssa Nicole 3MT1Alyssa Nicole BarrettoNo ratings yet
- Anatomy and Physiology of Female Reproductive System:: VaginaDocument5 pagesAnatomy and Physiology of Female Reproductive System:: VaginaphallicNo ratings yet
- LarynxDocument7 pagesLarynxMarsella Lulu AuliaNo ratings yet
- Digestive SystemDocument6 pagesDigestive SystemCeleste Atienza BawagNo ratings yet
- Anatomy and Physiology - Doc Onco WardDocument4 pagesAnatomy and Physiology - Doc Onco WardLory LynNo ratings yet
- Membranes and FasciaeDocument18 pagesMembranes and FasciaeBurak KıvırcıkNo ratings yet
- Complete Metal Crown & Bridge Preparation GuideDocument62 pagesComplete Metal Crown & Bridge Preparation GuidesrinivaskalluriNo ratings yet
- Presentation 1Document12 pagesPresentation 1Rizka FarahinNo ratings yet
- Jurnal Kesehatan Gigi: Hubungan Plak Gigi, Laju Aliran Saliva, Dan Viskositas Saliva Pada Anak Usia 6-9 TahunDocument4 pagesJurnal Kesehatan Gigi: Hubungan Plak Gigi, Laju Aliran Saliva, Dan Viskositas Saliva Pada Anak Usia 6-9 TahunFransiski HoNo ratings yet
- The Journal of Dermatology - 2022 - Sakamoto - T Cell Rich Perivascular Inflammation in A Case of Cutaneous VariantDocument2 pagesThe Journal of Dermatology - 2022 - Sakamoto - T Cell Rich Perivascular Inflammation in A Case of Cutaneous VariantKi IsoNo ratings yet
- Daftar PustakaDocument2 pagesDaftar PustakaNizarul HizomiNo ratings yet
- Excretion and Homeostasis: Maintaining the Body's Internal BalanceDocument11 pagesExcretion and Homeostasis: Maintaining the Body's Internal BalancestarcandypricessNo ratings yet