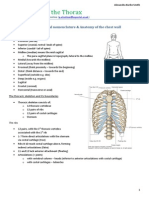
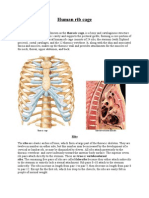
You might also like
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (121)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (588)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (400)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5794)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2259)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1090)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (344)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- Veterinary Anatomy Coloring BookDocument53 pagesVeterinary Anatomy Coloring BookJuliana Escudero100% (4)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (895)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- Sports Therapy For The Shoulder Return To The Sports PDFDocument240 pagesSports Therapy For The Shoulder Return To The Sports PDFGhinter MariusNo ratings yet
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (74)
- Practical Necropsy GuideDocument52 pagesPractical Necropsy GuideDulNo ratings yet
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- ABS-Anatomy of The ThoraxDocument52 pagesABS-Anatomy of The ThoraxAtti Tamas100% (1)
- ThompsonDocument95 pagesThompsonoooo100% (1)
- Pocket Book For PhysiotherapistDocument339 pagesPocket Book For PhysiotherapistEloisaNo ratings yet
- TSRA Review of Cardiothoracic PDFDocument298 pagesTSRA Review of Cardiothoracic PDFGaetano Di GiovanniNo ratings yet
- Skeletal SystemDocument13 pagesSkeletal SystemJohn Stephen CastilloNo ratings yet
- Moore's Clinically Oriented Anatomy 6th EdDocument4 pagesMoore's Clinically Oriented Anatomy 6th EdJohna Pauline Mandac67% (6)
- G4 Test 1Document5 pagesG4 Test 1Lindiwe GoweNo ratings yet
- What Is The Skeletal System?: Axial Skeleton AnatomyDocument2 pagesWhat Is The Skeletal System?: Axial Skeleton AnatomyJared RodriguezNo ratings yet
- Skeletal System Zoology ReportDocument15 pagesSkeletal System Zoology ReportHannah Nicole TomasNo ratings yet
- Sonoanatomy of The Spine A Comprehensive Scanning Protocol From Cervical To Sacral RegionDocument9 pagesSonoanatomy of The Spine A Comprehensive Scanning Protocol From Cervical To Sacral Regionfajarrudy qimindraNo ratings yet
- Surface Anat of ThoraxDocument42 pagesSurface Anat of ThoraxSyed NangyalNo ratings yet
- Bones and Joints Quiz Questions - Footprints-Science GCSE Science Animations and QuizzesDocument1 pageBones and Joints Quiz Questions - Footprints-Science GCSE Science Animations and Quizzescpcp6n7tc5No ratings yet
- Biology 3Document126 pagesBiology 3khaledNo ratings yet
- Respiratory Module Guidelines SapDocument5 pagesRespiratory Module Guidelines SapRabia OwaisNo ratings yet
- Interior Root WordsDocument3 pagesInterior Root WordsPaula McAuliffeNo ratings yet
- Human Rib CageDocument3 pagesHuman Rib CageAmerNo ratings yet
- Support Systems Grade 10Document51 pagesSupport Systems Grade 10Lance McGillNo ratings yet
- Score: 1: Emaciated 2: Thin 3: Good 4: Fat 5: ObeseDocument6 pagesScore: 1: Emaciated 2: Thin 3: Good 4: Fat 5: ObeseLaura RicoNo ratings yet
- Ch. 7 Student Packet (ANP 1)Document6 pagesCh. 7 Student Packet (ANP 1)Alex ZhangNo ratings yet
- 1.upper Limb and Thorax: Section 1Document6 pages1.upper Limb and Thorax: Section 1Labana LabanaNo ratings yet
- Osteology BonesDocument48 pagesOsteology BonesÂhmęd ĐoolaNo ratings yet
- Lab 7 Skeletal System PowerPointDocument19 pagesLab 7 Skeletal System PowerPointNguyen (Harry) Xuan HoangNo ratings yet
- Cervical RibDocument15 pagesCervical RibArko duttaNo ratings yet
- Thoracic Inlet Syndrome and Cervical RibDocument28 pagesThoracic Inlet Syndrome and Cervical RibRabin DasNo ratings yet
- Selina Solutions For Class 9 Biology Chapter 12 Skeleton Movement and LocomotionDocument9 pagesSelina Solutions For Class 9 Biology Chapter 12 Skeleton Movement and LocomotionAnonymous 404No ratings yet
- Specific Osteology: Skeleton Axiale - Cranium (Skull) - Truncus (Trunk)Document46 pagesSpecific Osteology: Skeleton Axiale - Cranium (Skull) - Truncus (Trunk)Defi Sofianti AnnoNo ratings yet
- Axial Body - Muscles (Head, Neck & Trunk)Document14 pagesAxial Body - Muscles (Head, Neck & Trunk)HaleyNo ratings yet