You might also like
- 12 Higher-Order and Subjective Aspects of PerceptionDocument16 pages12 Higher-Order and Subjective Aspects of PerceptionSilaxNo ratings yet
- Transferencia de CalorDocument5 pagesTransferencia de CalorSilax100% (1)
- Chapter 11 SYG Nephrin IrreC Family of Adhesion Proteins Mediate Asymmetric Cell-Cell Adhesion in DevelopmentDocument11 pagesChapter 11 SYG Nephrin IrreC Family of Adhesion Proteins Mediate Asymmetric Cell-Cell Adhesion in DevelopmentSilaxNo ratings yet
- A Comparison of The Carbon Balances of A Natural Lake and A Hydroelectric Reservoir in Northern SwedenDocument8 pagesA Comparison of The Carbon Balances of A Natural Lake and A Hydroelectric Reservoir in Northern SwedenSilaxNo ratings yet
- Front-Matter The Sticky SynapseDocument10 pagesFront-Matter The Sticky SynapseSilaxNo ratings yet
- 14 NAcetylaspartate and NAcetylaspartylglutamateDocument42 pages14 NAcetylaspartate and NAcetylaspartylglutamateSilaxNo ratings yet
- 24 Microtransplantation of Neurotransmitter Receptors From Cells To Xenopus Oocyte Membrane S New Procedure For Ion Channel StudiesDocument9 pages24 Microtransplantation of Neurotransmitter Receptors From Cells To Xenopus Oocyte Membrane S New Procedure For Ion Channel StudiesSilaxNo ratings yet
- 25 Reconstitution of Golgi Disassembly by Mitotic Xenopus Egg Extract in Semi-Intact MDCK CellsDocument9 pages25 Reconstitution of Golgi Disassembly by Mitotic Xenopus Egg Extract in Semi-Intact MDCK CellsSilaxNo ratings yet
- 13 Disorders of Amino Acid MetabolismDocument28 pages13 Disorders of Amino Acid MetabolismSilaxNo ratings yet
- Back MatterDocument13 pagesBack MatterSilaxNo ratings yet
- Low Molecular Weight Peptides and Their Role in the BrainDocument12 pagesLow Molecular Weight Peptides and Their Role in the BrainSilaxNo ratings yet
- 10 ChromatiN Assembly of DNA Templates Microinjected Into Xenopus OocytesDocument9 pages10 ChromatiN Assembly of DNA Templates Microinjected Into Xenopus OocytesSilaxNo ratings yet
- Ammonia Toxicity in the Central Nervous System: A Review of Key Mechanisms and EffectsDocument16 pagesAmmonia Toxicity in the Central Nervous System: A Review of Key Mechanisms and EffectsSilaxNo ratings yet
- 8 Taurine: S. S. Oja - P. SaransaariDocument52 pages8 Taurine: S. S. Oja - P. SaransaariSilaxNo ratings yet
- Amino Acids Role in Regulating Brain Volume During Osmotic ChangesDocument24 pagesAmino Acids Role in Regulating Brain Volume During Osmotic ChangesSilaxNo ratings yet
- 9 Neurobiology of DAmino AcidsDocument18 pages9 Neurobiology of DAmino AcidsSilaxNo ratings yet
- Unlock SGCh05Document11 pagesUnlock SGCh05SilaxNo ratings yet
- 11 Urea Cycle EnzymopathiesDocument12 pages11 Urea Cycle EnzymopathiesSilaxNo ratings yet
- 4 Aromatic Amino Acids in The BrainDocument40 pages4 Aromatic Amino Acids in The BrainSilaxNo ratings yet
- 6 Branched Chain Amino Acids (BCAAs) in BrainDocument16 pages6 Branched Chain Amino Acids (BCAAs) in BrainSilaxNo ratings yet
- 7 Sulfur Containing Amino AcidsDocument22 pages7 Sulfur Containing Amino AcidsSilaxNo ratings yet
- Table of ContentsDocument3 pagesTable of ContentsJorge Alberto Chavarría SacasaNo ratings yet
- Unlock SGCH04Document6 pagesUnlock SGCH04SilaxNo ratings yet
- 5 Arginine, Citrulline, and OrnithineDocument18 pages5 Arginine, Citrulline, and OrnithineSilaxNo ratings yet
- Brain Index: A Concise Guide to Key ConceptsDocument6 pagesBrain Index: A Concise Guide to Key ConceptsSilaxNo ratings yet
- How We Built The Quintessential Sentry Gun: (Updated Sept. 26th)Document11 pagesHow We Built The Quintessential Sentry Gun: (Updated Sept. 26th)SilaxNo ratings yet
- Motion Control HandbookDocument38 pagesMotion Control Handbookvincentttt100% (1)
- Unlock SGCh03Document9 pagesUnlock SGCh03SilaxNo ratings yet
- Unlock SGCh36Document10 pagesUnlock SGCh36SilaxNo ratings yet
- Unlock SGCh02Document13 pagesUnlock SGCh02SilaxNo ratings yet
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (890)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (587)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5794)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (399)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (344)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (73)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (265)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2219)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1090)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (119)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- Excel 2007 Lesson 7 QuizDocument5 pagesExcel 2007 Lesson 7 Quizdeep72No ratings yet
- ITTC Validation of ManoeuvringDocument11 pagesITTC Validation of ManoeuvringFelipe IlhaNo ratings yet
- HowTo Work With CR 90Document87 pagesHowTo Work With CR 90WagBezerraNo ratings yet
- Manual de Operacion de Bomba BlackmerDocument20 pagesManual de Operacion de Bomba BlackmerMorales EduardoNo ratings yet
- Indian Standards List As On Jan2009Document216 pagesIndian Standards List As On Jan2009Vasudeva Pavan VemuriNo ratings yet
- 000 200 1210 Guidelines For Minimum Deliverables 3 November 2011Document22 pages000 200 1210 Guidelines For Minimum Deliverables 3 November 2011Raul Bautista100% (1)
- Mathematics of Finance: Simple and Compound Interest FormulasDocument11 pagesMathematics of Finance: Simple and Compound Interest FormulasAshekin MahadiNo ratings yet
- User Mode I. System Support Processes: de Leon - Dolliente - Gayeta - Rondilla It201 - Platform Technology - TPDocument6 pagesUser Mode I. System Support Processes: de Leon - Dolliente - Gayeta - Rondilla It201 - Platform Technology - TPCariza DollienteNo ratings yet
- PDS - GulfSea Hydraulic AW Series-1Document2 pagesPDS - GulfSea Hydraulic AW Series-1Zaini YaakubNo ratings yet
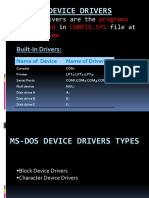
- Ms-Dos Device Drivers: Device Drivers Are The That in File atDocument13 pagesMs-Dos Device Drivers: Device Drivers Are The That in File atJass GillNo ratings yet
- PresiometroDocument25 pagesPresiometrojoseprepaNo ratings yet
- Design of Shaft Straightening MachineDocument58 pagesDesign of Shaft Straightening MachineChiragPhadkeNo ratings yet
- Answer Key Grade 6 - 2023Document7 pagesAnswer Key Grade 6 - 2023ALKHANSAA ELOBEIDYNo ratings yet
- Microstation V8I Accudraw Basics: Bentley Institute Course GuideDocument80 pagesMicrostation V8I Accudraw Basics: Bentley Institute Course Guideh_eijy2743No ratings yet
- Notifier Battery Calculations-ReadmeDocument11 pagesNotifier Battery Calculations-ReadmeJeanCarlosRiveroNo ratings yet
- Recommended Procedures For Internet-Based Connections Between Rths and Nmcs (VPN, Ipsec)Document38 pagesRecommended Procedures For Internet-Based Connections Between Rths and Nmcs (VPN, Ipsec)Crismaruc Maria-madalinaNo ratings yet
- Exponential & Logarithmic FunctionsDocument13 pagesExponential & Logarithmic FunctionsRahul SankaranNo ratings yet
- A Design and Analysis of A Morphing Hyper-Elliptic Cambered Span (HECS) WingDocument10 pagesA Design and Analysis of A Morphing Hyper-Elliptic Cambered Span (HECS) WingJEORJENo ratings yet
- DSD - Assignment 1 2018Document3 pagesDSD - Assignment 1 2018Naveen KumarNo ratings yet
- Camshaft Test SheetDocument4 pagesCamshaft Test SheetsughieantoNo ratings yet
- SPPID QuestionsDocument2 pagesSPPID Questionsvivek83% (12)
- ISO 8243 2013 Cigarettes - SamplingDocument18 pagesISO 8243 2013 Cigarettes - SamplingEko YuliantoNo ratings yet
- Employee performance factors analysis electronic companyDocument10 pagesEmployee performance factors analysis electronic companyAmrithaNo ratings yet
- Great Lakes PGDM Interview Access DetailsDocument1 pageGreat Lakes PGDM Interview Access DetailsJaswanth konkepudiNo ratings yet
- Affixation (Landscape)Document4 pagesAffixation (Landscape)difafalahudinNo ratings yet
- Verify scope acceptance of project deliverablesDocument5 pagesVerify scope acceptance of project deliverablesmypmpbooksNo ratings yet
- Geotehnical Engg. - AEE - CRPQsDocument48 pagesGeotehnical Engg. - AEE - CRPQsSureshKonamNo ratings yet
- Eltek PSR 327Document2 pagesEltek PSR 327fan liuNo ratings yet
- Cec 103. - Workshop Technology 1Document128 pagesCec 103. - Workshop Technology 1VietHungCao92% (13)
- Network Layer: Computer Networking: A Top Down ApproachDocument83 pagesNetwork Layer: Computer Networking: A Top Down ApproachMuhammad Bin ShehzadNo ratings yet