You might also like
- Earth First! Climbers Guild Vol. 2Document16 pagesEarth First! Climbers Guild Vol. 2CascadiaDefenders100% (2)
- Master Key Bonus - The Healing CodeDocument8 pagesMaster Key Bonus - The Healing CodeChanterelle Vasto100% (12)
- Better - Call - Saul - 4x10 WINNERDocument70 pagesBetter - Call - Saul - 4x10 WINNERMarcos Valdés GarridoNo ratings yet
- Pharmaceutical Microbiology - B.pharmDocument383 pagesPharmaceutical Microbiology - B.pharmkeyurNo ratings yet
- Old Fashioned Southern Tea CakesDocument2 pagesOld Fashioned Southern Tea CakesDB ScottNo ratings yet
- Innate ImmunityDocument44 pagesInnate ImmunityAhmed J AlhindaweNo ratings yet
- Major Emergencies in Power PlantDocument29 pagesMajor Emergencies in Power Plantsourav mahapatraNo ratings yet
- Music and Therapy Bun2Document17 pagesMusic and Therapy Bun2PanteaAlisaNo ratings yet
- Polymerase Chain ReactionDocument34 pagesPolymerase Chain Reactionmokshgoyal2597100% (4)
- Prevalence and Characterization of Shiga Toxin O157 H7 and Non O157 H7 Enterohemorrhagic Escherichia Coli Isolated From Different SourcesDocument10 pagesPrevalence and Characterization of Shiga Toxin O157 H7 and Non O157 H7 Enterohemorrhagic Escherichia Coli Isolated From Different SourcesSABA YOUNASNo ratings yet
- Molecular Characterization of Extended-Spectrum BetaLactamase (ESBLs) Genes in Pseudomonas Aeruginosa From Pregnant Women Attending A Tertiary Health Care Centre in Makurdi, Central NigeriaDocument7 pagesMolecular Characterization of Extended-Spectrum BetaLactamase (ESBLs) Genes in Pseudomonas Aeruginosa From Pregnant Women Attending A Tertiary Health Care Centre in Makurdi, Central NigeriaJASH MATHEWNo ratings yet
- Diarrhoeagenic Escherichia Coli in Mother Child Pairs in Ile-Ife, South Western NigeriaDocument9 pagesDiarrhoeagenic Escherichia Coli in Mother Child Pairs in Ile-Ife, South Western NigeriaTunde OdetoyinNo ratings yet
- MD 07024Document5 pagesMD 07024Ateeqa MalikNo ratings yet
- Pseudomonas Aeruginosa, 2013Document12 pagesPseudomonas Aeruginosa, 2013katiushikasNo ratings yet
- High Incidence of Multidrug Resistant Escherichia Coli Producing CTX-M-type ESBLs Colonizing The Intestine of Jordanian InfantsDocument8 pagesHigh Incidence of Multidrug Resistant Escherichia Coli Producing CTX-M-type ESBLs Colonizing The Intestine of Jordanian InfantsInternational Medical PublisherNo ratings yet
- 2014 Prevalence of Aeromonas Species Among Patients Attending General Hospital OwerriDocument10 pages2014 Prevalence of Aeromonas Species Among Patients Attending General Hospital OwerriJOHNKENNEDY NNODIMNo ratings yet
- 1 Soler 2008 FunctEcolDocument8 pages1 Soler 2008 FunctEcolalcedo76No ratings yet
- Antimicrobial Resistance Patterns of Diarrheagenic and Non - Diarrheagenic Escherichia Coli Isolates From Libyan ChildrenDocument8 pagesAntimicrobial Resistance Patterns of Diarrheagenic and Non - Diarrheagenic Escherichia Coli Isolates From Libyan ChildrenInternational Medical PublisherNo ratings yet
- Received: Accepted:: UjmrDocument5 pagesReceived: Accepted:: UjmrUMYU Journal of Microbiology Research (UJMR)No ratings yet
- 34 557 ElhaDocument8 pages34 557 ElhaYosi Dwi Saputro Part IINo ratings yet
- Virulence Factors of Enterococcus FaecalisDocument5 pagesVirulence Factors of Enterococcus FaecalisRatih PurnamaNo ratings yet
- Research ArticleDocument9 pagesResearch Articlepujari chaithanyaNo ratings yet
- Multi-Biochemical Test System For: Distinguishing Enteric and Other Gram-Negative BacilliDocument7 pagesMulti-Biochemical Test System For: Distinguishing Enteric and Other Gram-Negative BacilliSyazmin KhairuddinNo ratings yet
- Antibiotic Resistance Profile of Staphylococci From Clinical Sources Recovered From InfantsDocument7 pagesAntibiotic Resistance Profile of Staphylococci From Clinical Sources Recovered From InfantsCornius TerranovaNo ratings yet
- Klebsiella Pneumoniae: Trends in The Hospital and Community SettingsDocument5 pagesKlebsiella Pneumoniae: Trends in The Hospital and Community SettingskhanfahsNo ratings yet
- 5 Psae Egypt 2014Document9 pages5 Psae Egypt 2014Khoa Vi Sinh BVCRNo ratings yet
- Rate of Cultivable Subgingival Periodontopathogenic Bacteria in Chronic PeriodontitisDocument5 pagesRate of Cultivable Subgingival Periodontopathogenic Bacteria in Chronic PeriodontitisTahir AliNo ratings yet
- Possibe Association Beteween Mutldrug Resistenace and Isolate Origin With Som Virulence Factors of E Coli Strains Isolated From Infant FeacesDocument11 pagesPossibe Association Beteween Mutldrug Resistenace and Isolate Origin With Som Virulence Factors of E Coli Strains Isolated From Infant FeacesCarlos VazquezNo ratings yet
- Isolation and Characterization of Potential Probiotic Escherichia Coli Strains From Rat Faecal SamplesDocument7 pagesIsolation and Characterization of Potential Probiotic Escherichia Coli Strains From Rat Faecal Samplesada-adutzaNo ratings yet
- Begum 2013Document4 pagesBegum 2013WahyunitadotokaNo ratings yet
- Bacteriological and Pathological Investigation of Nasal Passage Infections of Chickens (Gallus Gallus)Document9 pagesBacteriological and Pathological Investigation of Nasal Passage Infections of Chickens (Gallus Gallus)Noman FarookNo ratings yet
- Abbas - 2016 - Bacteriological Study of Puerperal Sepsis in Al-Najaf CityDocument9 pagesAbbas - 2016 - Bacteriological Study of Puerperal Sepsis in Al-Najaf CityAbdu GodanaNo ratings yet
- Multiple-Antibiotic Resistance Mediated by Plasmids and Integrons in Uropathogenic Escherichia Coli and Klebsiella PneumoniaeDocument5 pagesMultiple-Antibiotic Resistance Mediated by Plasmids and Integrons in Uropathogenic Escherichia Coli and Klebsiella PneumoniaeProbioticsAnywhereNo ratings yet
- Discussion: and Camponotusfloridanus) and Various Niches (Focusing On The PAH Degrading Microbes) .InDocument10 pagesDiscussion: and Camponotusfloridanus) and Various Niches (Focusing On The PAH Degrading Microbes) .InfupaNo ratings yet
- 18 35 1 SMDocument10 pages18 35 1 SMhud_fNo ratings yet
- Rahul KambleDocument5 pagesRahul KambleAnonymous 6EHECZNo ratings yet
- Detect Mycoplasma Contamination Cell Cultures Sera PCRDocument7 pagesDetect Mycoplasma Contamination Cell Cultures Sera PCRmaniiiiiiiiNo ratings yet
- MelitusDocument6 pagesMelituswulan faddhylanNo ratings yet
- Coli With Gross and Histopathological Lesions in BroilerDocument8 pagesColi With Gross and Histopathological Lesions in BroilermaafiamNo ratings yet
- tmpC1AA TMPDocument3 pagestmpC1AA TMPFrontiersNo ratings yet
- AMR of Salmonella and Shig HararDocument6 pagesAMR of Salmonella and Shig Hararpurposef49No ratings yet
- 03-Av-Ho Thi Viet Thu (24-29) 003Document6 pages03-Av-Ho Thi Viet Thu (24-29) 003Quy NguyenNo ratings yet
- Aem 64 9 3336-3345 1998Document10 pagesAem 64 9 3336-3345 1998Juan ZurartNo ratings yet
- PDFDocument7 pagesPDFFredNo ratings yet
- Escherichia Coli Strains From Ostriches and TheirDocument9 pagesEscherichia Coli Strains From Ostriches and Theirali hendyNo ratings yet
- A Study of Various Isolates From Pus Sample With Their Antibiogram From JLN Hospital, AjmerDocument5 pagesA Study of Various Isolates From Pus Sample With Their Antibiogram From JLN Hospital, AjmerIOSRjournalNo ratings yet
- Isolation and Identification of Avibacterium Paragallinarum From Layer Chickens in Gazipur, BangladeshDocument3 pagesIsolation and Identification of Avibacterium Paragallinarum From Layer Chickens in Gazipur, BangladeshMd. Ehsanul Haque100% (1)
- Cold Ham 2013Document9 pagesCold Ham 2013Omar AguilarNo ratings yet
- E.coli O157 H7 Shiga ToxinDocument16 pagesE.coli O157 H7 Shiga ToxinNayshaNo ratings yet
- Prevalence of Streptococcus pneumoniae in Egyptian ChildrenDocument5 pagesPrevalence of Streptococcus pneumoniae in Egyptian Childrenelshaymaa Abdelsattar MohamedNo ratings yet
- Antibiogram of E. Coli Serotypes IsolatedDocument5 pagesAntibiogram of E. Coli Serotypes IsolatedFitriyana WinarnoNo ratings yet
- K.N.ravichitra, Et AlDocument5 pagesK.N.ravichitra, Et AldindaNo ratings yet
- Integrative Investigation Recognizes BHLH Transcription Factors As Contributing Factors For You To Parkinsons Ailment Threat ElementsnxtfqDocument2 pagesIntegrative Investigation Recognizes BHLH Transcription Factors As Contributing Factors For You To Parkinsons Ailment Threat Elementsnxtfqboatdance86No ratings yet
- Isolation of Total Aerobic and Pathogenic Bacteria From Table Eggs and ItsDocument9 pagesIsolation of Total Aerobic and Pathogenic Bacteria From Table Eggs and ItsnorsyahirahafiqahNo ratings yet
- Antibiotic Sensitivity Pattern and Plasmid Profile of Bacteria Isolated From Diabetic Ulcers in Mbano Metropolis, Imo State, Southeastern NigeriaDocument9 pagesAntibiotic Sensitivity Pattern and Plasmid Profile of Bacteria Isolated From Diabetic Ulcers in Mbano Metropolis, Imo State, Southeastern NigeriaUMYU Journal of Microbiology Research (UJMR)No ratings yet
- A Selective Medium For Fusobacterium SPPDocument4 pagesA Selective Medium For Fusobacterium SPPJood turkNo ratings yet
- tmp3F98 TMPDocument4 pagestmp3F98 TMPFrontiersNo ratings yet
- Microbewiki - Kenyon.edu Index - PHP Enterobacter CloacaeDocument4 pagesMicrobewiki - Kenyon.edu Index - PHP Enterobacter Cloacaehari.suryanto8417No ratings yet
- 2022 Serogroups and Genetic Diversity of Diarrheagenic Strains of Escherichia Coli A Retrospective StudyDocument8 pages2022 Serogroups and Genetic Diversity of Diarrheagenic Strains of Escherichia Coli A Retrospective StudyAnca MareNo ratings yet
- Psa 2Document6 pagesPsa 2Nurul Musfira MasdukiNo ratings yet
- Epidermidis Carrying Biofilm Formation GenesDocument5 pagesEpidermidis Carrying Biofilm Formation GenesLini MaliqisnayantiNo ratings yet
- Received: Accepted: 23/06/2019Document7 pagesReceived: Accepted: 23/06/2019UMYU Journal of Microbiology Research (UJMR)No ratings yet
- Hektoen Enteric Agar ProtocolDocument13 pagesHektoen Enteric Agar ProtocolYakisoba menNo ratings yet
- Isolation of Bacteria Associated With SPDocument8 pagesIsolation of Bacteria Associated With SPMr. Md. Imdadul HaqueNo ratings yet
- PDF 9Document7 pagesPDF 9Justy GuavaNo ratings yet
- Jurnal Kedokteran Dan Kesehatan IndonesiaDocument7 pagesJurnal Kedokteran Dan Kesehatan IndonesiaReffy AdhaNo ratings yet
- ESBL Producing MDR Urinary Pathogens in Nepali ChildrenDocument5 pagesESBL Producing MDR Urinary Pathogens in Nepali ChildrennawarajNo ratings yet
- JOURNALDocument28 pagesJOURNALskylarsavioNo ratings yet
- By:Hend Eldeeb: Complete The Following SentencesDocument6 pagesBy:Hend Eldeeb: Complete The Following SentencesAhmed J AlhindaweNo ratings yet
- Scanned by CamscannerDocument10 pagesScanned by CamscannerAhmed J AlhindaweNo ratings yet
- Scanned by CamscannerDocument11 pagesScanned by CamscannerAhmed J AlhindaweNo ratings yet
- Enterobacteriaceae Opportunistic PathogensDocument45 pagesEnterobacteriaceae Opportunistic PathogensAhmed J AlhindaweNo ratings yet
- Dissertation PDFDocument115 pagesDissertation PDFAhmed J AlhindaweNo ratings yet
- 787Document8 pages787Ahmed J AlhindaweNo ratings yet
- GrammarDocument118 pagesGrammarAhmed J AlhindaweNo ratings yet
- CamScanner Scans PDFs QuicklyDocument4 pagesCamScanner Scans PDFs QuicklyAhmed J AlhindaweNo ratings yet
- 55052Document14 pages55052Ahmed J AlhindaweNo ratings yet
- Identification of Enterobacter SPP by 16SrRNAgene Sequencing in Basrah Province IraqDocument14 pagesIdentification of Enterobacter SPP by 16SrRNAgene Sequencing in Basrah Province IraqAhmed J AlhindaweNo ratings yet
- 55439Document13 pages55439Ahmed J AlhindaweNo ratings yet
- PlasmidDocument15 pagesPlasmidndiugbegbeNo ratings yet
- Seminar of Cell Culture TechniquesDocument55 pagesSeminar of Cell Culture TechniquesAhmed J AlhindaweNo ratings yet
- Escherichia ColiDocument32 pagesEscherichia ColiAhmed J AlhindaweNo ratings yet
- (O, H, Vi) Free ImmunoprotectiveDocument9 pages(O, H, Vi) Free ImmunoprotectiveAhmed J AlhindaweNo ratings yet
- Bilal 1Document48 pagesBilal 1Ahmed J AlhindaweNo ratings yet
- History of Animal Tissue Culture and Natural Surroundings For Animal CellDocument16 pagesHistory of Animal Tissue Culture and Natural Surroundings For Animal CellAhmed J AlhindaweNo ratings yet
- DNA-based meat speciation techniquesDocument4 pagesDNA-based meat speciation techniquesAhmed J AlhindaweNo ratings yet
- 48721Document33 pages48721Ahmed J AlhindaweNo ratings yet
- Rapid IsolationDocument2 pagesRapid IsolationAhmed J AlhindaweNo ratings yet
- Meat TestDocument3 pagesMeat TestAhmed J AlhindaweNo ratings yet
- 32721Document33 pages32721prasadbheemNo ratings yet
- 032 Icetd2012 D10046Document7 pages032 Icetd2012 D10046Ahmed J AlhindaweNo ratings yet
- Histone Variants: Are They Functionally Heterogeneous?: David T BrownDocument6 pagesHistone Variants: Are They Functionally Heterogeneous?: David T Brownnath211No ratings yet
- 1Document2 pages1Ahmed J AlhindaweNo ratings yet
- Windows 8 NoticeDocument1 pageWindows 8 NoticeAhmed J AlhindaweNo ratings yet
- DNA Structure and Central DogmaDocument5 pagesDNA Structure and Central DogmaAhmed J AlhindaweNo ratings yet
- Project 3 - Gas Turbine Tutorial - ChemCad Draft - 01 15 2020Document58 pagesProject 3 - Gas Turbine Tutorial - ChemCad Draft - 01 15 2020Daniel Andres Canro CalderónNo ratings yet
- Chemistry in Everyday LifeDocument33 pagesChemistry in Everyday LifeArya StarkNo ratings yet
- Thai Occupational Safety, Health and Environment Act B.E. 2554 (A.d. 2011)Document32 pagesThai Occupational Safety, Health and Environment Act B.E. 2554 (A.d. 2011)DibbaSotaNanaNo ratings yet
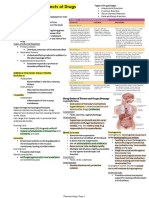
- CHAPTER 3: Toxic Effects of Drugs: Pharmacology Page 1Document1 pageCHAPTER 3: Toxic Effects of Drugs: Pharmacology Page 1Gabriel GonzagaNo ratings yet
- E06 Power Cable Joints and Terminations v1Document20 pagesE06 Power Cable Joints and Terminations v1Anorld WalkerNo ratings yet
- Indian Medical Tourism Industry: A Pathway For The Healthy Future of IndiaDocument13 pagesIndian Medical Tourism Industry: A Pathway For The Healthy Future of IndiaPranjal MaluNo ratings yet
- SITE. Defining Project Goals & ObjectivesDocument1 pageSITE. Defining Project Goals & Objectivescoach balaNo ratings yet
- RV Failure: Pathophysiology, Diagnosis and TreatmentDocument16 pagesRV Failure: Pathophysiology, Diagnosis and TreatmentRizky Regia TriseynesyaNo ratings yet
- Substrates P400 enDocument111 pagesSubstrates P400 enLince WijoyoNo ratings yet
- Color Me VeganDocument272 pagesColor Me VeganBela Luísa Novo100% (1)
- STC Bec Wir Plu D 0209 01 PDFDocument10 pagesSTC Bec Wir Plu D 0209 01 PDFAdel MorsyNo ratings yet
- Tap ChangerDocument5 pagesTap ChangerRavindra AngalNo ratings yet
- RESOLUTION 2022-08 Denial of Setback VarianceDocument5 pagesRESOLUTION 2022-08 Denial of Setback VarianceMatt McKinneyNo ratings yet
- April 12th Test PDFDocument32 pagesApril 12th Test PDFهخه •No ratings yet
- CapecitabineDocument8 pagesCapecitabineLisa MarieNo ratings yet
- Implementation of Swachh Bharat in MysoreDocument13 pagesImplementation of Swachh Bharat in MysoreDipyaman ChoudhuryNo ratings yet
- BiostastisticDocument2 pagesBiostastisticاحمد ماجد زبنNo ratings yet
- Comparing Food with Comparative AdjectivesDocument4 pagesComparing Food with Comparative AdjectivesLuisito GonzalezNo ratings yet
- 02 - Purulent Inflammatory DiseaseDocument60 pages02 - Purulent Inflammatory Diseaseshekhawatyuvraj051No ratings yet
- Deliver VALUE and RENEWABLE FUTUREDocument30 pagesDeliver VALUE and RENEWABLE FUTUREAlexandru SuciuNo ratings yet
- 3M Bio-Acoustic Simulator User Manual (053-012 Rev D)Document21 pages3M Bio-Acoustic Simulator User Manual (053-012 Rev D)rodrigocornejo81No ratings yet
- Holy Childhood High School Chemistry 601 Topic: Electronic StructureDocument6 pagesHoly Childhood High School Chemistry 601 Topic: Electronic StructureBriana BartonNo ratings yet
- Nihms 223615 PDFDocument15 pagesNihms 223615 PDFElay PedrosoNo ratings yet