You might also like
- Feline Asthma: A Pathophysiologic Basis of TherapyDocument5 pagesFeline Asthma: A Pathophysiologic Basis of TherapyEnnur NufianNo ratings yet
- Tibio-Tarsal Luxation and Its Management in A Dog: Ayyappan, S., M. Shiju Simon, B. C. Das and R. Suresh KumarDocument4 pagesTibio-Tarsal Luxation and Its Management in A Dog: Ayyappan, S., M. Shiju Simon, B. C. Das and R. Suresh KumarEnnur NufianNo ratings yet
- Perbandingan Kadar Elektrolit Serum Pascaenterektomi Ekstensif 75 % Pada Anjing Yang Diterapi Dengan LaktoferinDocument7 pagesPerbandingan Kadar Elektrolit Serum Pascaenterektomi Ekstensif 75 % Pada Anjing Yang Diterapi Dengan LaktoferinEnnur NufianNo ratings yet
- Pengaruh Lama Thawing Terhadap Kualitas Semen Beku Sapi Limousin Dan BrahmanDocument5 pagesPengaruh Lama Thawing Terhadap Kualitas Semen Beku Sapi Limousin Dan BrahmanEnnur NufianNo ratings yet
- Summary Profile For Enzootic Bovine Leukosis (Ebl) : Vetsurveillance@defra - Gsi.gov - UkDocument2 pagesSummary Profile For Enzootic Bovine Leukosis (Ebl) : Vetsurveillance@defra - Gsi.gov - UkEnnur NufianNo ratings yet
- Rimsa12 18-EDocument14 pagesRimsa12 18-EEnnur NufianNo ratings yet
- Pengembangan Teknik Elisa Untuk Mendeteksi Toksin Alfa ClostridiumnovyiDocument6 pagesPengembangan Teknik Elisa Untuk Mendeteksi Toksin Alfa ClostridiumnovyiEnnur NufianNo ratings yet
- Daftar Pustaka: - 1987. "Accounting and - Harvard Business School PressDocument8 pagesDaftar Pustaka: - 1987. "Accounting and - Harvard Business School PressEnnur NufianNo ratings yet
- Book Revue: Review A de LivreDocument1 pageBook Revue: Review A de LivreEnnur NufianNo ratings yet
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5794)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (895)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (588)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (400)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2259)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (74)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (345)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (121)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- Amazonas Nov Dic18Document100 pagesAmazonas Nov Dic18Sebastian GaunaNo ratings yet
- Migration in BirdsDocument29 pagesMigration in BirdsGEETA MOHAN100% (1)
- Snakes of South AfricaDocument576 pagesSnakes of South AfricaPedro V. M. ChacónNo ratings yet
- Hygiene Que. BSS 1) What Are Insects?Document2 pagesHygiene Que. BSS 1) What Are Insects?Amaresh JhaNo ratings yet
- Tanner Method and AndrogynyDocument16 pagesTanner Method and AndrogynyNur Fatmah SaidNo ratings yet
- Coastal Crabs A Guide To The Crabs of New Zealand 2015Document55 pagesCoastal Crabs A Guide To The Crabs of New Zealand 2015Carlos GarciaNo ratings yet
- Ministerio de Turismo: Zarpo Zarpo Zarpo Zarpo Zarpo Zarpo Zarpo Zarpo Zarpo ZarpoDocument10 pagesMinisterio de Turismo: Zarpo Zarpo Zarpo Zarpo Zarpo Zarpo Zarpo Zarpo Zarpo Zarpocarina vrnturaNo ratings yet
- Std09 I SSS EM 1 PDFDocument114 pagesStd09 I SSS EM 1 PDFSindhu GunaNo ratings yet
- Biology - The Genetics of Parenthood AnalysisDocument2 pagesBiology - The Genetics of Parenthood Analysislanichung100% (5)
- Analogy (Advanced) - Reasoning Shortcuts: Reasoning For Bank Exams 21 CommentsDocument6 pagesAnalogy (Advanced) - Reasoning Shortcuts: Reasoning For Bank Exams 21 CommentsSai Sree CharanNo ratings yet
- Histopathology ETASDocument89 pagesHistopathology ETASDr.Tawheed75% (4)
- Gas Exchange in Animals: We Will Be Studying The Diversity of Adaptations For This Process in Four Animal GroupsDocument42 pagesGas Exchange in Animals: We Will Be Studying The Diversity of Adaptations For This Process in Four Animal GroupsMiss IqbalNo ratings yet
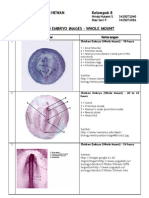
- Chick Embryo WMDocument6 pagesChick Embryo WMdhyan_ajjah67% (3)
- Full Basic Psychopharmacology For Counselors and Psychotherapists 2Nd Edition Sinacola Test Bank PDFDocument25 pagesFull Basic Psychopharmacology For Counselors and Psychotherapists 2Nd Edition Sinacola Test Bank PDFwilliam.ely879100% (16)
- Chapter 3 - Chicken Anatomy and Physiology: Figure 3.1 - The Internal Organs of The Female ChickenDocument3 pagesChapter 3 - Chicken Anatomy and Physiology: Figure 3.1 - The Internal Organs of The Female ChickenwijakesumaNo ratings yet
- Bryophytes of Central Province Sri LankaDocument6 pagesBryophytes of Central Province Sri LankaAshen NirodyaNo ratings yet
- The Ontogenetic Basic of Human AnatomyDocument18 pagesThe Ontogenetic Basic of Human AnatomymeseniaNo ratings yet
- Fish and BreedingDocument404 pagesFish and BreedingJohn100% (1)
- Flyer Manuale - CON WATERMARK PAGINE SINGOLEDocument6 pagesFlyer Manuale - CON WATERMARK PAGINE SINGOLEjscmtNo ratings yet
- Code: VT802-KIT: DS WS1 WS2Document2 pagesCode: VT802-KIT: DS WS1 WS2Sep NeufNo ratings yet
- Formicidae PDFDocument3,127 pagesFormicidae PDFCindy GarciaNo ratings yet
- Carpophilus Keys For Identification of Carpophilus PDFDocument41 pagesCarpophilus Keys For Identification of Carpophilus PDFErica Livea100% (3)
- PETA PEMIKIRAN (I-Think)Document4 pagesPETA PEMIKIRAN (I-Think)Fazwan YusoffNo ratings yet
- Invertebrate ZoologyDocument16 pagesInvertebrate ZoologyRenz Francis SasaNo ratings yet
- Echinodermata Notes - 2023Document2 pagesEchinodermata Notes - 2023BHEKUMUSA MASEKONo ratings yet
- You Will Hear Peter Walsh Being Interviewed For A Job. Listen and Choose The Correct Answer For Each QuestionDocument8 pagesYou Will Hear Peter Walsh Being Interviewed For A Job. Listen and Choose The Correct Answer For Each QuestionHiền NhiNo ratings yet
- The Use of Insects in Forensic Investigations - An Overview On The Scope of Forensic EntomologyDocument4 pagesThe Use of Insects in Forensic Investigations - An Overview On The Scope of Forensic EntomologyLight100% (1)
- 4HB0 Revision NotesDocument4 pages4HB0 Revision NotesTamer A. DakkakNo ratings yet
- Avifaunal Diversity of Bega Watershed, Prosperidad, Agusan Del Sur, PhilippinesDocument16 pagesAvifaunal Diversity of Bega Watershed, Prosperidad, Agusan Del Sur, PhilippinesInternational Network For Natural SciencesNo ratings yet
- Pharyngeal Arch Derivatives Chart!Document5 pagesPharyngeal Arch Derivatives Chart!Suphadetch Leung100% (1)