You might also like
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (119)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (265)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (399)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (587)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5794)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2219)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1090)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (344)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (894)
- Grow Food Grow Hope Toolkit Updated To Share OnlineDocument35 pagesGrow Food Grow Hope Toolkit Updated To Share OnlineWilmington College Grow Food Grow Hope100% (1)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (73)
- Landscape 3Document64 pagesLandscape 3lakshmi akshaya maddiNo ratings yet
- (Tony Sato) Ariocarpus. Hand Book PDFDocument93 pages(Tony Sato) Ariocarpus. Hand Book PDFOsmarUrielReyesAscencioNo ratings yet
- Roses and Their Care PDFDocument2 pagesRoses and Their Care PDFut.testbox7243No ratings yet
- 2010HeweziArabidopsis Spermidine Synthase Is Targeted by An Effector Protein of The Cyst Nematode Heterodera Schachtii-1Document3 pages2010HeweziArabidopsis Spermidine Synthase Is Targeted by An Effector Protein of The Cyst Nematode Heterodera Schachtii-1naelscribdNo ratings yet
- Lattice ManualDocument146 pagesLattice ManualnaelscribdNo ratings yet
- F 3951 EBO The Origin of Parasitism Gene in Nematodes Evolutionary Analysis Thro - PDF 5303Document14 pagesF 3951 EBO The Origin of Parasitism Gene in Nematodes Evolutionary Analysis Thro - PDF 5303naelscribdNo ratings yet
- 2013AtkinsonIdentification of Genes Involved in The Response of Arabidopsis To Simultaneous Biotic and Abiotic StressesDocument14 pages2013AtkinsonIdentification of Genes Involved in The Response of Arabidopsis To Simultaneous Biotic and Abiotic StressesnaelscribdNo ratings yet
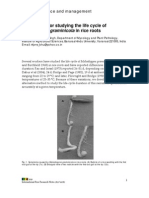
- A Technique For Studying The Life Cycle of Meloidogyne Graminicola in Rice RootsDocument3 pagesA Technique For Studying The Life Cycle of Meloidogyne Graminicola in Rice RootsGrace CañasNo ratings yet
- Ihsan Prot Lipid ArticleDocument9 pagesIhsan Prot Lipid ArticlenaelscribdNo ratings yet
- 2010 Nematodes Manual ENGLISHDocument93 pages2010 Nematodes Manual ENGLISHnaelscribdNo ratings yet
- Metabolic Profiling Reveals Local and Systemic ResponsesDocument14 pagesMetabolic Profiling Reveals Local and Systemic ResponsesnaelscribdNo ratings yet
- Cyst Extraction From SoilDocument4 pagesCyst Extraction From SoilnaelscribdNo ratings yet
- Identification of Novel Human Genes Evolutionarily Conserved inDocument12 pagesIdentification of Novel Human Genes Evolutionarily Conserved innaelscribdNo ratings yet
- Large-Scale Sequencing Reveals 21U-RNAs and Additional MicroRNAs and Endogenous SiRNAs in C. ElegansDocument15 pagesLarge-Scale Sequencing Reveals 21U-RNAs and Additional MicroRNAs and Endogenous SiRNAs in C. ElegansnaelscribdNo ratings yet
- Composition and Dynamics of The Caenorhabditis Elegans Early Embryonic TranscriptomeDocument12 pagesComposition and Dynamics of The Caenorhabditis Elegans Early Embryonic TranscriptomenaelscribdNo ratings yet
- NGC Product Reg List - Peru 20240206Document3 pagesNGC Product Reg List - Peru 20240206jichumpitazfNo ratings yet
- Instant Download Test Bank For Chemical Dependency A Systems Approach 4 e 4th Edition C Aaron Mcneece Diana M Dinitto PDF ScribdDocument32 pagesInstant Download Test Bank For Chemical Dependency A Systems Approach 4 e 4th Edition C Aaron Mcneece Diana M Dinitto PDF Scribdjasonzunigapdonjzbmws100% (11)
- The Nutrient Content of Paspalum Atratum Grass Associated With Macroptilium Lathyroides Legume Inoculated With Rhizobium Through The Application of Molybdenum and Phosphorus FertilizersDocument5 pagesThe Nutrient Content of Paspalum Atratum Grass Associated With Macroptilium Lathyroides Legume Inoculated With Rhizobium Through The Application of Molybdenum and Phosphorus FertilizersMamta AgarwalNo ratings yet
- Mohan BrinjalDocument25 pagesMohan BrinjalHARISH LALNo ratings yet
- YaraDocument5 pagesYaraAna StarciucNo ratings yet
- Plant Parts and Their FunctionDocument10 pagesPlant Parts and Their FunctionSetyo NugrohoNo ratings yet
- Yellow Poinciana Fact SheetDocument3 pagesYellow Poinciana Fact SheetNandhitha RajarathnamNo ratings yet
- Anatomy of Plant Anmol SirDocument24 pagesAnatomy of Plant Anmol SirKhushi PathakNo ratings yet
- Earth and Life Science: Plant Cells vs Animal CellsDocument48 pagesEarth and Life Science: Plant Cells vs Animal CellssailorNo ratings yet
- Green Fingers Student WorksheetDocument4 pagesGreen Fingers Student Worksheetionela -simona PopescuNo ratings yet
- Selection of Plants in LandscapeDocument32 pagesSelection of Plants in LandscapeSakshi RawatNo ratings yet
- Model Bankable Project On Bamboo Based Agroforestry: National Bank For Agriculture and Rural DevelopmentDocument24 pagesModel Bankable Project On Bamboo Based Agroforestry: National Bank For Agriculture and Rural DevelopmentBalakrishna GopinathNo ratings yet
- Green Manure in CoconutDocument4 pagesGreen Manure in CoconutNandakumar PalaparambilNo ratings yet
- Laboratory Activity No. 2Document10 pagesLaboratory Activity No. 2neilandreiruba1011No ratings yet
- SatyamDocument13 pagesSatyamnegisrishti27No ratings yet
- Intercropping Cereals and Grain Legumes - A Farmers PerspectiveDocument2 pagesIntercropping Cereals and Grain Legumes - A Farmers PerspectivesedianpoNo ratings yet
- On Killing A TreeDocument4 pagesOn Killing A TreejockimdNo ratings yet
- Med-RM Bot SP-1 AnswersDocument7 pagesMed-RM Bot SP-1 Answerskrish masterjeeNo ratings yet
- Catalogo AlforexDocument16 pagesCatalogo AlforexXavi SudNo ratings yet
- Seed Structure and Function: Nature's Blueprint for Plant GrowthDocument5 pagesSeed Structure and Function: Nature's Blueprint for Plant GrowthJohn AplerhNo ratings yet
- Seed Formation and DevelopmentDocument12 pagesSeed Formation and DevelopmentSabir Ahamed90% (20)
- Soal Bahasa Inggris Minggu Ke 2Document4 pagesSoal Bahasa Inggris Minggu Ke 2zaharNo ratings yet
- Identification and Morphological Characterization of Pathogens Infecting Melia Dubia in Tamil NaduDocument8 pagesIdentification and Morphological Characterization of Pathogens Infecting Melia Dubia in Tamil NaduSp SarathyNo ratings yet
- Seed PrimingDocument4 pagesSeed PrimingbigbangNo ratings yet
- Ethiopia livestock forage guide on pasture crops, conservationDocument35 pagesEthiopia livestock forage guide on pasture crops, conservationabraha gebruNo ratings yet
- Tropical Woods 1955 - 101-103Document99 pagesTropical Woods 1955 - 101-103HozaifaNo ratings yet