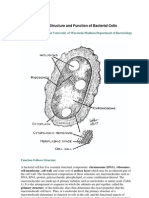
You might also like
- Formation and Fate of Cell OrganellesFrom EverandFormation and Fate of Cell OrganellesKatherine Brehme WarrenNo ratings yet
- Introduction of BacteriaDocument15 pagesIntroduction of BacteriaAsif IqbalNo ratings yet
- Differences between Prokaryotic and Eukaryotic CellsDocument122 pagesDifferences between Prokaryotic and Eukaryotic CellsAdithya NanuvalaNo ratings yet
- A Guide To Wastewater MicrobiologyDocument14 pagesA Guide To Wastewater Microbiologyabduyassin8014No ratings yet
- AP Biology Bacteria and VirusesDocument51 pagesAP Biology Bacteria and VirusesMichael TsaiNo ratings yet
- Bacterial Cell StructureDocument6 pagesBacterial Cell StructureCasey StuartNo ratings yet
- Unit 2 (1) 2Document11 pagesUnit 2 (1) 2Johnieer Bassem MoferdNo ratings yet
- Ultrastructure of Bacteria & EndosporeDocument8 pagesUltrastructure of Bacteria & EndosporeSubrata KunduNo ratings yet
- MicrobiologyDocument8 pagesMicrobiologyKeishaAaliyahNo ratings yet
- Microbiology and Immunology Department: Bacterial GrowthDocument8 pagesMicrobiology and Immunology Department: Bacterial GrowthAbir OmarNo ratings yet
- LECTURE 3 ProkaryotesDocument33 pagesLECTURE 3 ProkaryotesNicole MandyNo ratings yet
- Parker 2001Document6 pagesParker 2001Chinasa EkejiubaNo ratings yet
- Prokaryotic Cells Are Also CalledDocument25 pagesProkaryotic Cells Are Also CalledArdo RamdhaniNo ratings yet
- Mod 2Document24 pagesMod 2The GreatNo ratings yet
- The CellDocument5 pagesThe CellAfida Razuna AveNo ratings yet
- LECT 2 ProkaryotesDocument30 pagesLECT 2 ProkaryotesjudynakwebwaNo ratings yet
- Bacterial ClassificationDocument5 pagesBacterial ClassificationShai KateNo ratings yet
- Bahan Ajar Bio Sel Bab 1-4Document92 pagesBahan Ajar Bio Sel Bab 1-4Maychel Arientawira JohannisNo ratings yet
- Prokaryotic and Eukaryotic CellsDocument21 pagesProkaryotic and Eukaryotic CellskenNo ratings yet
- Bacterial Cell Wall TypesDocument7 pagesBacterial Cell Wall Typess.khan9211rediffmail.comNo ratings yet
- Bacterial MorphologyDocument11 pagesBacterial Morphologydavid_physiologyNo ratings yet
- Presentation 2Document54 pagesPresentation 2hunter zoneNo ratings yet
- BiochemistryDocument531 pagesBiochemistrySiss Thwae100% (1)
- Bab 6 Sruktur Dan Fungsi Sel Bakteria (I)Document9 pagesBab 6 Sruktur Dan Fungsi Sel Bakteria (I)Shakira AmiraNo ratings yet
- FAQ in MicrobiologyDocument117 pagesFAQ in MicrobiologyVenkatesh Arumugam100% (1)
- Differences Between Prokaryotic and Eukaryotic CellsDocument8 pagesDifferences Between Prokaryotic and Eukaryotic CellsMarcy BoralNo ratings yet
- Biochemistry The Chemical Reactions of Living Cells 2d Ed Vols 1 2 David E. MetzlerDocument1,977 pagesBiochemistry The Chemical Reactions of Living Cells 2d Ed Vols 1 2 David E. MetzlerEvelyn Biscari100% (7)
- Microbiology Lecture on Bacterial Morphology and StainingDocument7 pagesMicrobiology Lecture on Bacterial Morphology and StainingAhmed AliNo ratings yet
- Bacteria Cell StructureDocument4 pagesBacteria Cell StructureReyhan Audian Dwi PutraNo ratings yet
- The Bacterial Cell EnvelopeDocument19 pagesThe Bacterial Cell EnvelopetheiasitNo ratings yet
- Essay 3Document3 pagesEssay 3Tri HardiyantiNo ratings yet
- BIL Exam 3 NotesDocument16 pagesBIL Exam 3 NotesCraig ContrerasNo ratings yet
- The Plasma (Cytoplasmic) MembraneDocument6 pagesThe Plasma (Cytoplasmic) Membraneprism1702No ratings yet
- BIO2107 - Tutorial 1 Discussion PointsDocument5 pagesBIO2107 - Tutorial 1 Discussion PointsAjay Sookraj RamgolamNo ratings yet
- For Ntse CellDocument73 pagesFor Ntse CellPrakhar RajNo ratings yet
- Cell and Molecular Biology Concepts and Experiments 7th Edition Karp Solutions Manual DownloadDocument49 pagesCell and Molecular Biology Concepts and Experiments 7th Edition Karp Solutions Manual DownloadBen Flott100% (24)
- Structure & Functions of Prokaryotic Cell. Bacteriophages.: Lecturer: - Sergei V. RedkozubovDocument54 pagesStructure & Functions of Prokaryotic Cell. Bacteriophages.: Lecturer: - Sergei V. RedkozubovJustineNo ratings yet
- Eukaryote BiologyDocument9 pagesEukaryote BiologyKrizzy EstoceNo ratings yet
- Overview of Microbial LifeDocument9 pagesOverview of Microbial LifeQuan ThieuNo ratings yet
- Differences Between Prokaryotic and Eukaryotic Cells (39Document49 pagesDifferences Between Prokaryotic and Eukaryotic Cells (39Jeanine CristobalNo ratings yet
- The History of LifeThe Eukaryota Include The Organisms That Most People Are Most Familiar WithDocument5 pagesThe History of LifeThe Eukaryota Include The Organisms That Most People Are Most Familiar WithbjoteroNo ratings yet
- The Cell: Anabolism CatabolismDocument14 pagesThe Cell: Anabolism CatabolismHashim GhazoNo ratings yet
- Bacteria Cell Structure 300Document13 pagesBacteria Cell Structure 300Success OlamideNo ratings yet
- 1 - Give An Illustrated Account of The Fine Staracture of A Plant Cell and Describe The Function of The Diffrent Cell OrgnagesDocument19 pages1 - Give An Illustrated Account of The Fine Staracture of A Plant Cell and Describe The Function of The Diffrent Cell OrgnagesMD shah KhanNo ratings yet
- Pys PDFDocument5 pagesPys PDFMD shah KhanNo ratings yet
- Bacteria and Archea: Structural Ad Functional AdaptationsDocument9 pagesBacteria and Archea: Structural Ad Functional AdaptationshomamunfatNo ratings yet
- 1.3 Bacterial MorphologyDocument4 pages1.3 Bacterial MorphologyJosh Miguel BorromeoNo ratings yet
- The Bacterial Cell: Structures of Prokaryotes and Differences from EukaryotesDocument3 pagesThe Bacterial Cell: Structures of Prokaryotes and Differences from EukaryotesanojanNo ratings yet
- Cell The Unit of Life: 16 June 2022 03:52Document19 pagesCell The Unit of Life: 16 June 2022 03:52Dhanalakshmi LakshmiNo ratings yet
- Bacterial Cell StructureDocument9 pagesBacterial Cell StructureManWol JangNo ratings yet
- ArchaeDocument6 pagesArchaesudamsahu197No ratings yet
- Inclusion 1.3Document6 pagesInclusion 1.3debbyblingsNo ratings yet
- The essential base of Cell BiologyDocument16 pagesThe essential base of Cell BiologyTambudzai RazyNo ratings yet
- 1.the Bacterial Cell Wall StructureDocument10 pages1.the Bacterial Cell Wall StructureMark Jhon MoradaNo ratings yet
- Prokaryotic and Eukaryotic Cells 3.1 CellDocument9 pagesProkaryotic and Eukaryotic Cells 3.1 CellJosebeth CairoNo ratings yet
- COT EnglishDocument4 pagesCOT EnglishTypie ZapNo ratings yet
- Marksmanship: Subject: III. Definition of TermsDocument16 pagesMarksmanship: Subject: III. Definition of TermsAmber EbayaNo ratings yet
- Technical File D13-MH, MG IMO Tier 11 GLDocument18 pagesTechnical File D13-MH, MG IMO Tier 11 GLsfsdffdsdfsdfsdfNo ratings yet
- CV Abdalla Ali Hashish-Nursing Specialist.Document3 pagesCV Abdalla Ali Hashish-Nursing Specialist.Abdalla Ali HashishNo ratings yet
- Bitcoin MambaDocument17 pagesBitcoin MambaJean MilenixNo ratings yet
- Zhihua Yao - Dignaga and The 4 Types of Perception (JIP 04)Document24 pagesZhihua Yao - Dignaga and The 4 Types of Perception (JIP 04)Carlos Caicedo-Russi100% (1)
- The Emperor Jones: What's Inside in ContextDocument27 pagesThe Emperor Jones: What's Inside in ContextHarshvardhan RaiNo ratings yet
- CIGB B164 Erosion InterneDocument163 pagesCIGB B164 Erosion InterneJonathan ColeNo ratings yet
- Aquafine Optivenn Series Data SheetDocument8 pagesAquafine Optivenn Series Data SheetKenz ZhouNo ratings yet
- No.6 Role-Of-Child-Health-NurseDocument8 pagesNo.6 Role-Of-Child-Health-NursePawan BatthNo ratings yet
- Plant Air Centrifugal Compressors: Turbo-Air Series Featuring Oil-Free AirDocument20 pagesPlant Air Centrifugal Compressors: Turbo-Air Series Featuring Oil-Free AirSharad KokateNo ratings yet
- Desarme Del ConvertidorpdfDocument7 pagesDesarme Del ConvertidorpdfDiego Orlando Santos BuitragoNo ratings yet
- The Polynesians: Task1: ReadingDocument10 pagesThe Polynesians: Task1: ReadingHəşim MəmmədovNo ratings yet
- K Series Parts List - 091228Document25 pagesK Series Parts List - 091228AstraluxNo ratings yet
- Health Information System Developmen T (Medical Records)Document21 pagesHealth Information System Developmen T (Medical Records)skidz137217100% (10)
- Castel - From Dangerousness To RiskDocument10 pagesCastel - From Dangerousness To Riskregmatar100% (2)
- Application Programming InterfaceDocument12 pagesApplication Programming InterfacesorinproiecteNo ratings yet
- Ch. 7 - Audit Reports CA Study NotesDocument3 pagesCh. 7 - Audit Reports CA Study NotesUnpredictable TalentNo ratings yet
- Kastanakis 2014Document8 pagesKastanakis 2014Andreea Georgiana MocanuNo ratings yet
- Startups Helping - India Go GreenDocument13 pagesStartups Helping - India Go Greensimran kNo ratings yet
- Mechanical Questions & AnswersDocument161 pagesMechanical Questions & AnswersTobaNo ratings yet
- 3 Steel Grating Catalogue 2010 - SERIES 1 PDFDocument6 pages3 Steel Grating Catalogue 2010 - SERIES 1 PDFPablo MatrakaNo ratings yet
- E PortfolioDocument76 pagesE PortfolioMAGALLON ANDREWNo ratings yet
- Product Catalog 2016Document84 pagesProduct Catalog 2016Sauro GordiniNo ratings yet
- Assessing Eyes NCM 103 ChecklistDocument7 pagesAssessing Eyes NCM 103 ChecklistNicole NipasNo ratings yet
- Korba - BAH Online Temp MonitoringDocument7 pagesKorba - BAH Online Temp Monitoringrama jenaNo ratings yet
- 2.0 - SITHKOP002 - Plan and Cost Basic Menus Student GuideDocument92 pages2.0 - SITHKOP002 - Plan and Cost Basic Menus Student Guidebash qwertNo ratings yet
- Biomechanics of Advanced Tennis: January 2003Document7 pagesBiomechanics of Advanced Tennis: January 2003Katrien BalNo ratings yet
- Raychem Price ListDocument48 pagesRaychem Price ListramshivvermaNo ratings yet
- Brochure - Truemax Concrete Pump Truck Mounted TP25M4Document16 pagesBrochure - Truemax Concrete Pump Truck Mounted TP25M4RizkiRamadhanNo ratings yet