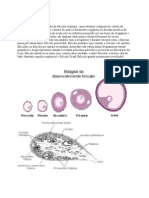
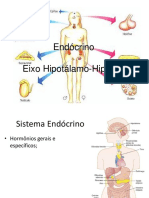
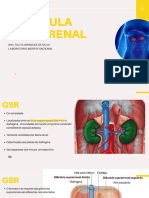
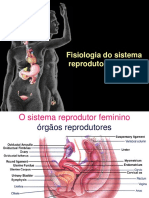
You might also like
- Degeneração TesticularDocument14 pagesDegeneração TesticularGabrielle MouraNo ratings yet
- Departamento de Ciências de Saúd1Document13 pagesDepartamento de Ciências de Saúd1Milagre FernandesNo ratings yet
- EMBRIOLOGIADocument22 pagesEMBRIOLOGIAAna LídiaNo ratings yet
- FOLICULOGÊNESE Ntit (Recovered)Document8 pagesFOLICULOGÊNESE Ntit (Recovered)Simone Salvador BoeneNo ratings yet
- Morfofisiologia MasculinoDocument8 pagesMorfofisiologia MasculinoAdolfo SilvaNo ratings yet
- Rel Bio 12Document6 pagesRel Bio 12Luísa Braz TeixeiraNo ratings yet
- Apostila Andrologico Biotran - PRNDocument27 pagesApostila Andrologico Biotran - PRNHenry Chinaski100% (1)
- Sistema Reprodutor MasculinoDocument9 pagesSistema Reprodutor MasculinoAssistência Social ingazeiraNo ratings yet
- Trabalho de EspermatogeneseDocument8 pagesTrabalho de EspermatogeneseScarlett DuarteNo ratings yet
- Histologia e EmbriologiaDocument3 pagesHistologia e EmbriologiaJuliana MoraesNo ratings yet
- Biologia Básica II UnifatecieDocument32 pagesBiologia Básica II UnifatecieAline LopesNo ratings yet
- EmbriologiaDocument9 pagesEmbriologiacb_penatrujilloNo ratings yet
- Embriologia ResumoDocument3 pagesEmbriologia ResumoKarine EstrelaNo ratings yet
- Biologia 12.º AnoDocument65 pagesBiologia 12.º AnoPaula Amado100% (1)
- Roteiro Teorico Pratico PDFDocument16 pagesRoteiro Teorico Pratico PDFBeathryz LethycyaNo ratings yet
- Trabalho de BiologiaDocument13 pagesTrabalho de BiologiawaldenizaNo ratings yet
- Questões de Vestibulares - Embriologia (Gabaritada)Document6 pagesQuestões de Vestibulares - Embriologia (Gabaritada)netoalvirubroNo ratings yet
- Relatório 1Document14 pagesRelatório 1Alexandra BalçaNo ratings yet
- Sistema Reprodutor Masculino e GametogêneseDocument2 pagesSistema Reprodutor Masculino e GametogêneseDanielle MariaNo ratings yet
- AULA 1 - Introdução Ao Estudo Da Morfologia e EmbriologiaDocument76 pagesAULA 1 - Introdução Ao Estudo Da Morfologia e EmbriologiaMarcelle Gonçalves de MirandaNo ratings yet
- Embriologia - Luciana TalianoDocument39 pagesEmbriologia - Luciana TalianoLuciana TalianoNo ratings yet
- EmbriologiaDocument6 pagesEmbriologiaFelipe SantosNo ratings yet
- Funcões Reprodutoras e Hormonais MasculinasDocument6 pagesFuncões Reprodutoras e Hormonais MasculinasZanon de Oliveira, AgendamentoNo ratings yet
- Gametogênese - ResumoDocument3 pagesGametogênese - Resumofabiodevs100% (2)
- GametogeneseDocument6 pagesGametogeneseLetícia BoniniNo ratings yet
- Embriologia de Todos Os SistemasDocument44 pagesEmbriologia de Todos Os SistemasKeyla SilvaNo ratings yet
- ReprodutoresDocument50 pagesReprodutoresJoão Feca MussoqueNo ratings yet
- Null 2Document12 pagesNull 2adaoNo ratings yet
- Questionario Av2 2021Document5 pagesQuestionario Av2 2021Letycia Fernandes100% (1)
- Relatório Biologia 12ºano - Observação de EspermatozóidesDocument10 pagesRelatório Biologia 12ºano - Observação de EspermatozóidesMarisa Milhano80% (5)
- (EMBRIOLOGIA - RESUMO) EspermatogêneseDocument37 pages(EMBRIOLOGIA - RESUMO) EspermatogêneseCaroline MilheiroNo ratings yet
- 0.MA - Elemento Textual - Morfologia HumanaDocument109 pages0.MA - Elemento Textual - Morfologia HumanaChristiane BarbiNo ratings yet
- Estudo Dirigido Embriologia 1 CópiaDocument8 pagesEstudo Dirigido Embriologia 1 CópiamarianacndcNo ratings yet
- EmbriologiaDocument6 pagesEmbriologiaLetícia BoniniNo ratings yet
- Apostila Anatomia FloralDocument13 pagesApostila Anatomia FloralIsabella CapillaNo ratings yet
- Atividade Divina 1Document4 pagesAtividade Divina 1DannyMendesNo ratings yet
- Estudo Dirigido 1 - EmbriologiaDocument6 pagesEstudo Dirigido 1 - EmbriologiaCarla Caroline Alves De LimaNo ratings yet
- Embriologia 2011 - GabaritoDocument2 pagesEmbriologia 2011 - GabaritoIvaneaVasquesCruzNo ratings yet
- Histologia e EmbriologiaDocument16 pagesHistologia e Embriologialulu.helena2806No ratings yet
- Estudo Dirigido EMBRIOLOGIADocument10 pagesEstudo Dirigido EMBRIOLOGIAValeria OliveiraNo ratings yet
- Material G2021Document42 pagesMaterial G2021Amina OssufoNo ratings yet
- Embriologia Humana - Parte 1Document30 pagesEmbriologia Humana - Parte 1Yam VictorNo ratings yet
- Apostila - EmbriologiaDocument96 pagesApostila - EmbriologiaGuilhermeSantosNo ratings yet
- Biologia 1,2.3.4.5Document17 pagesBiologia 1,2.3.4.5JoaoNo ratings yet
- Prova de Embriologia - Andressa PintoDocument3 pagesProva de Embriologia - Andressa PintoAndressa AzevedoNo ratings yet
- Fisiologia v2 Semana09Document7 pagesFisiologia v2 Semana09nrogernNo ratings yet
- Ud I - Sexualidade Humana PDFDocument80 pagesUd I - Sexualidade Humana PDFPedro HenriqueNo ratings yet
- Anat.e Fisiol - Sistema Reprodutor Masculino 19pgDocument19 pagesAnat.e Fisiol - Sistema Reprodutor Masculino 19pgderek0% (1)
- 06 - Anatomofisiologia Do Sistema ReprodutorDocument21 pages06 - Anatomofisiologia Do Sistema ReprodutorCristiano PereiraNo ratings yet
- Embriologia Descomplicada - Flavio Chame BarretoDocument142 pagesEmbriologia Descomplicada - Flavio Chame BarretoMario MelloNo ratings yet
- Estudo Dirigido Embrio IDocument4 pagesEstudo Dirigido Embrio IdaienescamposNo ratings yet
- CITOLOGIA EMBRIOlogiaDocument4 pagesCITOLOGIA EMBRIOlogiaekarina265No ratings yet
- Aula 1 - Segmentação e BlastulaDocument50 pagesAula 1 - Segmentação e BlastulalahfmchNo ratings yet
- Lâmina 45Document3 pagesLâmina 45Lídia Imaculada TeixeiraNo ratings yet
- Resumão GlaDocument16 pagesResumão Gladanie0949No ratings yet
- Gametogénese e Regulação HormonalDocument53 pagesGametogénese e Regulação HormonalMaria SantosNo ratings yet
- 1 Desen Embrio e Difer SexualDocument17 pages1 Desen Embrio e Difer SexualChácara Arte SuaveNo ratings yet
- Triagem NeonatalDocument13 pagesTriagem Neonatalアメイジング グレイスNo ratings yet
- Aula 2 - Manejo Reprodutivo de Novilhas LeiteirasDocument45 pagesAula 2 - Manejo Reprodutivo de Novilhas LeiteirasNayara MartinsNo ratings yet
- Carte Chakra Couleur A4 Generale Contours Po PDFDocument1 pageCarte Chakra Couleur A4 Generale Contours Po PDFRenoir FonteasNo ratings yet
- TiroideDocument6 pagesTiroideSusana Correia100% (1)
- Reprodução - Fisiologia e Endocrinologia Do Ciclo EstralDocument13 pagesReprodução - Fisiologia e Endocrinologia Do Ciclo EstralRebeca MoreiraNo ratings yet
- BOLFER - HipotireoidismoDocument7 pagesBOLFER - HipotireoidismoVictoriabertoziNo ratings yet
- Sistema EndócrinoDocument33 pagesSistema EndócrinoGabriela MirandaNo ratings yet
- Aula 4 - Sistema TegumentarDocument39 pagesAula 4 - Sistema Tegumentargabykbm100% (1)
- REPRODUÇÃO em Medicina VeterináriaDocument54 pagesREPRODUÇÃO em Medicina Veterináriajamilesada100% (11)
- Apostila Módulo 3 - Exames Laboratoriais 2.0Document26 pagesApostila Módulo 3 - Exames Laboratoriais 2.0Eva Vitorino100% (3)
- Eixo Hipotálamo-Hipófise FarmDocument23 pagesEixo Hipotálamo-Hipófise FarmRafaela MacanhaoNo ratings yet
- Ratos VirtuaisDocument5 pagesRatos VirtuaisAnderson CarvalhoNo ratings yet
- Sistema Endocrino, - Urinario - NervosoDocument4 pagesSistema Endocrino, - Urinario - NervosoStyleziNNo ratings yet
- Fisiopatologia Da TireoideDocument3 pagesFisiopatologia Da TireoidePamella Carollyne De Souza Noronha MaiaNo ratings yet
- Glândula AdrenalDocument2 pagesGlândula AdrenalGabriela Ferst De RéNo ratings yet
- AVALIAÇÃO PRESENCIAL Anatomia Humana IIDocument2 pagesAVALIAÇÃO PRESENCIAL Anatomia Humana IIRennie Dourado PlateroNo ratings yet
- Téorica 2 Glandulas EndocrinasDocument43 pagesTéorica 2 Glandulas EndocrinasDanielle FeioNo ratings yet
- Aula Distúrbios Da Glândula TireóideDocument20 pagesAula Distúrbios Da Glândula TireóideSamuel Moreno100% (1)
- Menstruação PDFDocument7 pagesMenstruação PDFTayla OliveiraNo ratings yet
- Fisiologia EndócrinaDocument55 pagesFisiologia EndócrinaTalita RomanichenNo ratings yet
- Anatomia Da Glandula Supra RenalDocument33 pagesAnatomia Da Glandula Supra RenalAna Clara MoreiraNo ratings yet
- Aula 4 - Sistema Reprodutor MasculinoDocument3 pagesAula 4 - Sistema Reprodutor MasculinoVivi GlNo ratings yet
- Hipofise - HistologiaDocument8 pagesHipofise - HistologiaLetícia Alves PrudenteNo ratings yet
- NEUROPEPTÍDEOSDocument38 pagesNEUROPEPTÍDEOSJessica Lee100% (3)
- Aula 2 - Fisiologia Do Aparelho Reprodutor FemininoDocument23 pagesAula 2 - Fisiologia Do Aparelho Reprodutor FemininoSuellennobreNo ratings yet
- O Sistema Endócrino e Os ChakrasDocument3 pagesO Sistema Endócrino e Os ChakrasFernando CRNo ratings yet
- Sistema Genital FemininoDocument44 pagesSistema Genital FemininoSara CasteloNo ratings yet
- Os Chakras e As Glândulas e Órgãos CorrespondentesDocument2 pagesOs Chakras e As Glândulas e Órgãos CorrespondentesjaqueNo ratings yet
- Tireoidite de HaschimotoDocument7 pagesTireoidite de HaschimotoCamila SoaresNo ratings yet
- Hipotireoidismo Felino CongênitoDocument7 pagesHipotireoidismo Felino CongênitoMariana RNo ratings yet