You might also like
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (121)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (588)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (399)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5794)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2259)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (344)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1090)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (895)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (73)
- Benjamin, Walter - The Arcades ProjectDocument1,122 pagesBenjamin, Walter - The Arcades ProjectKostasSpinoza100% (1)
- The Nervous System PDFDocument3 pagesThe Nervous System PDFPerry SinNo ratings yet
- The SOMATID - A Pleomorphic, Ultra-Microsopic Subcellular Living and Reproducing EntityDocument55 pagesThe SOMATID - A Pleomorphic, Ultra-Microsopic Subcellular Living and Reproducing EntityMarkMadMunkiNo ratings yet
- 10 Rules For Parents of Defiant Kids With Adhd SecuredDocument1 page10 Rules For Parents of Defiant Kids With Adhd SecuredMário JorgeNo ratings yet
- Steel-Concrete Composite Coupling Beams - Behavior and DesignDocument11 pagesSteel-Concrete Composite Coupling Beams - Behavior and DesigncyrusnasiraiNo ratings yet
- Health As A Multifactorial PhenomenonDocument12 pagesHealth As A Multifactorial PhenomenonContessa Gabriel100% (1)
- Emma Holiday PediatricsDocument78 pagesEmma Holiday PediatricsRyan Franz100% (1)
- Cytogenetics, FISH and Molecular Testing in Hematologic Malignancies, 2008, p.334 PDFDocument334 pagesCytogenetics, FISH and Molecular Testing in Hematologic Malignancies, 2008, p.334 PDFJorge Santos100% (1)
- Limfoma Maligna FinalDocument50 pagesLimfoma Maligna FinalAulia Ayu PuspitaNo ratings yet
- A New Concept For The Contact at The Interface of Steel-Concrete Composite BeamsDocument23 pagesA New Concept For The Contact at The Interface of Steel-Concrete Composite BeamsTuấn Anh PhạmNo ratings yet
- Identification of Heavy Metal Pollutants Using Multivariate Analysis and Effects of Land Uses On Their Accumulation in Urban Soils in Beijing, ChinaDocument9 pagesIdentification of Heavy Metal Pollutants Using Multivariate Analysis and Effects of Land Uses On Their Accumulation in Urban Soils in Beijing, ChinaTuấn Anh PhạmNo ratings yet
- The Impact of Education, Income, and Mortality On Fertility in JamaicaDocument14 pagesThe Impact of Education, Income, and Mortality On Fertility in JamaicaTuấn Anh PhạmNo ratings yet
- A New Concept For The Contact at The Interface of Steel-Concrete Composite BeamsDocument23 pagesA New Concept For The Contact at The Interface of Steel-Concrete Composite BeamsTuấn Anh PhạmNo ratings yet
- IEEE Transactions on Power Electronics Volume 12 Issue 6 1997 [Doi 10.1109_63.641500] Yim-Shu Lee; Bo-Tao Lin -- Adding Active Clamping and Soft Switching to Boost-flyback Single-stage Isolated Power-factor-corrected Power SuppliesDocument11 pagesIEEE Transactions on Power Electronics Volume 12 Issue 6 1997 [Doi 10.1109_63.641500] Yim-Shu Lee; Bo-Tao Lin -- Adding Active Clamping and Soft Switching to Boost-flyback Single-stage Isolated Power-factor-corrected Power SuppliesTuấn Anh PhạmNo ratings yet
- IET Power Electronics Volume 4 Issue 9 2011 [Doi 10.1049_iet-Pel.2010.0363] Jung, J.-h.; Ahmed, S. -- Flyback Converter With Novel Active Clamp Control and Secondary Side Post Regulator for Low Standby Power Consumption Under HighDocument10 pagesIET Power Electronics Volume 4 Issue 9 2011 [Doi 10.1049_iet-Pel.2010.0363] Jung, J.-h.; Ahmed, S. -- Flyback Converter With Novel Active Clamp Control and Secondary Side Post Regulator for Low Standby Power Consumption Under HighTuấn Anh PhạmNo ratings yet
- A Chitosan-Gelatin Blend As A Coating For Fish PattiesDocument9 pagesA Chitosan-Gelatin Blend As A Coating For Fish PattiesTuấn Anh PhạmNo ratings yet
- IEEE Transactions on Power Electronics Volume 26 Issue 5 2011 [Doi 10.1109_tpel.2009.2022827] Lakshminarasamma, N.; Masihuzzaman, M.; Ramanarayanan, V. -- Steady-State Stability of Current-Mode Active-Clamp ZVS DC–DC ConvertersDocument10 pagesIEEE Transactions on Power Electronics Volume 26 Issue 5 2011 [Doi 10.1109_tpel.2009.2022827] Lakshminarasamma, N.; Masihuzzaman, M.; Ramanarayanan, V. -- Steady-State Stability of Current-Mode Active-Clamp ZVS DC–DC ConvertersTuấn Anh PhạmNo ratings yet
- In Viva Assay of Protein Synthesizing Capacity of Escherichia Coli From Slowly Growing Chemostat CulturesDocument14 pagesIn Viva Assay of Protein Synthesizing Capacity of Escherichia Coli From Slowly Growing Chemostat CulturesTuấn Anh PhạmNo ratings yet
- Nature's Nocturnal Services - Light Pollution As A Non-Recognised Challenge For Ecosystem Services Research and ManagementDocument5 pagesNature's Nocturnal Services - Light Pollution As A Non-Recognised Challenge For Ecosystem Services Research and ManagementTuấn Anh PhạmNo ratings yet
- IEEE Transactions on Power Electronics Volume 25 Issue 7 2010 [Doi 10.1109_tpel.2010.2042302] Junming Zhang; Xiucheng Huang; Xinke Wu; Zhaoming Qian -- A High Efficiency Flyback Converter With New Active Clamp TechniqueDocument11 pagesIEEE Transactions on Power Electronics Volume 25 Issue 7 2010 [Doi 10.1109_tpel.2010.2042302] Junming Zhang; Xiucheng Huang; Xinke Wu; Zhaoming Qian -- A High Efficiency Flyback Converter With New Active Clamp TechniqueTuấn Anh PhạmNo ratings yet
- An Isolated Interleaved Active-Clamp ZVT Flyback-Boost Converter With Coupled InductorsDocument9 pagesAn Isolated Interleaved Active-Clamp ZVT Flyback-Boost Converter With Coupled InductorsTuấn Anh PhạmNo ratings yet
- 20 de Luyen Thi Vao 10 4062Document32 pages20 de Luyen Thi Vao 10 4062cocghe2No ratings yet
- Giôùi Thieäu Chung Veà Nhieäm VuïDocument73 pagesGiôùi Thieäu Chung Veà Nhieäm VuïTuấn Anh PhạmNo ratings yet
- Higher-Order Cone-Pseudoconvex, Quasiconvex and Other Related Functions in Vector OptimizationDocument18 pagesHigher-Order Cone-Pseudoconvex, Quasiconvex and Other Related Functions in Vector OptimizationTuấn Anh PhạmNo ratings yet
- In Vitro Protein Refolding by Chromatographic ProceduresDocument10 pagesIn Vitro Protein Refolding by Chromatographic ProceduresTuấn Anh PhạmNo ratings yet
- Tong Hop Ngu Phap Tieng Anh Luyen Thi Dai HocDocument177 pagesTong Hop Ngu Phap Tieng Anh Luyen Thi Dai HocTuấn Anh PhạmNo ratings yet
- A Model For The Effects of Psychological Pricing in Gabor-Granger Price StudiesDocument24 pagesA Model For The Effects of Psychological Pricing in Gabor-Granger Price StudiesTuấn Anh PhạmNo ratings yet
- 1500 Test C 1538Document429 pages1500 Test C 1538Tuấn Anh PhạmNo ratings yet
- Nature's Nocturnal Services - Light Pollution As A Non-Recognised Challenge For Ecosystem Services Research and ManagementDocument5 pagesNature's Nocturnal Services - Light Pollution As A Non-Recognised Challenge For Ecosystem Services Research and ManagementTuấn Anh PhạmNo ratings yet
- Test 1 I. Give The Correct Form of Verbs GivenDocument21 pagesTest 1 I. Give The Correct Form of Verbs GivenTuấn Anh PhạmNo ratings yet
- Current Status of Technical Protein RefoldingDocument10 pagesCurrent Status of Technical Protein RefoldingTuấn Anh PhạmNo ratings yet
- Measuring Population HealthDocument29 pagesMeasuring Population HealthTuấn Anh PhạmNo ratings yet
- The Economics of Global Light PollutionDocument8 pagesThe Economics of Global Light PollutionTuấn Anh PhạmNo ratings yet
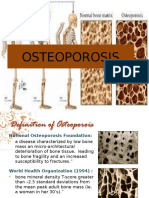
- OsteoporosisDocument13 pagesOsteoporosisMimi Suhaini SudinNo ratings yet
- Definition and Criteria For CKDDocument2 pagesDefinition and Criteria For CKDalejandraNo ratings yet
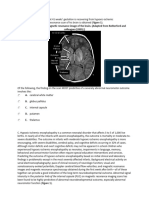
- Alzheimer's Disease: 2017 Facts and FiguresDocument88 pagesAlzheimer's Disease: 2017 Facts and FiguresGina SmithNo ratings yet
- Pre - & Suffix For HematologyDocument2 pagesPre - & Suffix For HematologyFrieda Cayabyab0% (1)
- Early HumanDocument4 pagesEarly HumanPeem JatabutNo ratings yet
- Four Major Drug TargetsDocument34 pagesFour Major Drug TargetsFlowerNo ratings yet
- Flow Cytometry: Surface Markers and Beyond: Methods of Allergy and ImmunologyDocument10 pagesFlow Cytometry: Surface Markers and Beyond: Methods of Allergy and ImmunologyMunir AliNo ratings yet
- Focal Adhesion Dynamics in Cellular Function and DiseaseDocument19 pagesFocal Adhesion Dynamics in Cellular Function and DiseaseEly Huaman PorrasNo ratings yet
- Chem Lab Final Exam Notes 2Document7 pagesChem Lab Final Exam Notes 2Jaira Emmarina100% (1)
- Chemical Carcinogenesis I and Ii: Michael Lea 2016Document31 pagesChemical Carcinogenesis I and Ii: Michael Lea 2016aasiyaNo ratings yet
- Nurse Writing 003 OET Practice Letter by PASS OETDocument26 pagesNurse Writing 003 OET Practice Letter by PASS OETNajad Salahudeen50% (2)
- Biology Paper 1 MsDocument20 pagesBiology Paper 1 Mssohaib0% (1)
- Diagnosis Power PointDocument26 pagesDiagnosis Power PointGlenn ChavezNo ratings yet
- Biological Impact of Feeding Rats With A Genetically Modified-Based DietDocument11 pagesBiological Impact of Feeding Rats With A Genetically Modified-Based DietJesús Rafael Méndez NateraNo ratings yet
- IgG Vs IgEDocument2 pagesIgG Vs IgEvyasakandarpNo ratings yet
- Sample Write UpDocument2 pagesSample Write UpStarr NewmanNo ratings yet
- NeoPREP 2011Document342 pagesNeoPREP 2011johnbarrow641No ratings yet
- Genetic EngineeringDocument14 pagesGenetic Engineering4L Anisha SieudassNo ratings yet
- WL7 Abstracts BookDocument1,514 pagesWL7 Abstracts BookJaison Enrique Torres PachecoNo ratings yet
- Addiction AND Upregulation BUISSON 2002Document7 pagesAddiction AND Upregulation BUISSON 2002Andra SilvaNo ratings yet
- Can Genes Predict Athletic Performance?Document2 pagesCan Genes Predict Athletic Performance?Channy PhamNo ratings yet
- Structure of ADC-68, a novel carbapenem-hydrolyzing class C extended-spectrum β-lactamase isolated from Acinetobacter baumanniiDocument14 pagesStructure of ADC-68, a novel carbapenem-hydrolyzing class C extended-spectrum β-lactamase isolated from Acinetobacter baumanniigeerthi vasanNo ratings yet
- Biology SL P2Document8 pagesBiology SL P2KenanNo ratings yet





















































![IEEE Transactions on Power Electronics Volume 12 Issue 6 1997 [Doi 10.1109_63.641500] Yim-Shu Lee; Bo-Tao Lin -- Adding Active Clamping and Soft Switching to Boost-flyback Single-stage Isolated Power-factor-corrected Power Supplies](https://imgv2-1-f.scribdassets.com/img/document/220209507/149x198/ca926c349c/1398397011?v=1)
![IET Power Electronics Volume 4 Issue 9 2011 [Doi 10.1049_iet-Pel.2010.0363] Jung, J.-h.; Ahmed, S. -- Flyback Converter With Novel Active Clamp Control and Secondary Side Post Regulator for Low Standby Power Consumption Under High](https://imgv2-1-f.scribdassets.com/img/document/220209509/149x198/8baf096670/1429428106?v=1)

![IEEE Transactions on Power Electronics Volume 26 Issue 5 2011 [Doi 10.1109_tpel.2009.2022827] Lakshminarasamma, N.; Masihuzzaman, M.; Ramanarayanan, V. -- Steady-State Stability of Current-Mode Active-Clamp ZVS DC–DC Converters](https://imgv2-2-f.scribdassets.com/img/document/220209510/149x198/51f107be20/1398397125?v=1)


![IEEE Transactions on Power Electronics Volume 25 Issue 7 2010 [Doi 10.1109_tpel.2010.2042302] Junming Zhang; Xiucheng Huang; Xinke Wu; Zhaoming Qian -- A High Efficiency Flyback Converter With New Active Clamp Technique](https://imgv2-1-f.scribdassets.com/img/document/220209508/149x198/263184eeda/1398397017?v=1)