Professional Documents
Culture Documents
El Ácido Desoxirribonucleico
Uploaded by
Guillermo GalvánCopyright
Available Formats
Share this document
Did you find this document useful?
Is this content inappropriate?
Report this DocumentCopyright:
Available Formats
El Ácido Desoxirribonucleico
Uploaded by
Guillermo GalvánCopyright:
Available Formats
El cido desoxirribonucleico, abreviado como ADN, es un cido nucleico que contiene instrucciones genticas
usadas en el desarrollo y funcionamiento de todos los organismos vivos conocidos y algunos virus, y es
responsable de su transmisin hereditaria. El papel principal de la molcula de ADN es el almacenamiento a
largo plazo de informacin. Muchas veces, el ADN es comparado con un plano o una receta, o un cdigo, ya que
contiene las instrucciones necesarias para construir otros componentes de las clulas, como las protenas y las
molculas de ARN. Los segmentos de ADN que llevan esta informacin gentica son llamados genes, pero las
otras secuencias de ADN tienen propsitos estructurales o toman parte en la regulacin del uso de esta
informacin gentica.
Desde el punto de vista qumico, el ADN es un polmero de nucletidos, es decir, un polinucletido. Un polmero
es un compuesto formado por muchas unidades simples conectadas entre s, como si fuera un largo tren
formado por vagones. En el ADN, cada vagn es un nucletido, y cada nucletido, a su vez, est formado por un
azcar (la desoxirribosa), una base nitrogenada (que puede ser adeninaA, timinaT, citosinaC o
guaninaG) y un grupo fosfato que acta como enganche de cada vagn con el siguiente. Lo que distingue a un
vagn (nucletido) de otro es, entonces, la base nitrogenada, y por ello la secuencia del ADN se especifica
nombrando slo la secuencia de sus bases. La disposicin secuencial de estas cuatro bases a lo largo de la
cadena (el ordenamiento de los cuatro tipos de vagones a lo largo de todo el tren) es la que codifica la
informacin gentica: por ejemplo, una secuencia de ADN puede ser ATGCTAGATCGC... En los organismos vivos,
el ADN se presenta como una doble cadena de nucletidos, en la que las dos hebras estn unidas entre s por
unas conexiones denominadas puentes de hidrgeno.1
Para que la informacin que contiene el ADN pueda ser utilizada por la maquinaria celular, debe copiarse en
primer lugar en unos trenes de nucletidos, ms cortos y con unas unidades diferentes, llamados ARN. Las
molculas de ARN se copian exactamente del ADN mediante un proceso denominado transcripcin. Una vez
procesadas en el ncleo celular, las molculas de ARN pueden salir al citoplasma para su utilizacin posterior. La
informacin contenida en el ARN se interpreta usando el cdigo gentico, que especifica la secuencia de los
aminocidos de las protenas, segn una correspondencia de un triplete de nucletidos (codn) para cada
aminocido. Esto es, la informacin gentica (esencialmente: qu protenas se van a producir en cada momento
del ciclo de vida de una clula) se halla codificada en las secuencias de nucletidos del ADN y debe traducirse
para poder funcionar. Tal traduccin se realiza usando el cdigo gentico a modo de diccionario. El diccionario
"secuencia de nucletido-secuencia de aminocidos" permite el ensamblado de largas cadenas de aminocidos
(las protenas) en el citoplasma de la clula. Por ejemplo, en el caso de la secuencia de ADN indicada antes
(ATGCTAGATCGC...), la ARN polimerasa utilizara como molde la cadena complementaria de dicha secuencia de
ADN (que sera TAC-GAT-CTA-GCG-...) para transcribir una molcula de ARNm que se leera AUG-CUA-GAU-CGC-
... ; el ARNm resultante, utilizando el cdigo gentico, se traducira como la secuencia de aminocidos
metionina-leucina-cido asprtico-arginina-...
Las secuencias de ADN que constituyen la unidad fundamental, fsica y funcional de la herencia se denominan
genes. Cada gen contiene una parte que se transcribe a ARN y otra que se encarga de definir cundo y dnde
deben expresarse. La informacin contenida en los genes (gentica) se emplea para generar ARN y protenas,
que son los componentes bsicos de las clulas, los "ladrillos" que se utilizan para la construccin de los
orgnulos u organelos celulares, entre otras funciones.
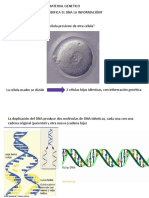
Dentro de las clulas, el ADN est organizado en estructuras llamadas cromosomas que, durante el ciclo celular,
se duplican antes de que la clula se divida. Los organismos eucariotas (por ejemplo, animales, plantas, y
hongos) almacenan la mayor parte de su ADN dentro del ncleo celular y una mnima parte en elementos
celulares llamados mitocondrias, y en los plastos y los centros organizadores de microtbulos o centrolos, en
caso de tenerlos; los organismos procariotas (bacterias y arqueas) lo almacenan en el citoplasma de la clula, y,
por ltimo, los virus ADN lo hacen en el interior de la cpsida de naturaleza proteica. Existen multitud de
protenas, como por ejemplo las histonas y los factores de transcripcin, que se unen al ADN dotndolo de una
estructura tridimensional determinada y regulando su expresin. Los factores de transcripcin reconocen
secuencias reguladoras del ADN y especifican la pauta de transcripcin de los genes. El material gentico
completo de una dotacin cromosmica se denomina genoma y, con pequeas variaciones, es caracterstico de
cada especie.
Propiedades fsicas y qumicas
Estructura qumica del ADN: dos cadenas de nucletidos conectadas mediante puentes de hidrgeno, que
aparecen como lneas punteadas.
El ADN es un largo polmero formado por unidades repetitivas, los nucletidos.
18
19
Una doble cadena de ADN
mide de 22 a 26 angstroms (2,2 a 2,6 nanmetros) de ancho, y una unidad (un nucletido) mide 3,3 (0,33 nm)
de largo.
20
Aunque cada unidad individual que se repite es muy pequea, los polmeros de ADN pueden
ser molculasenormes que contienen millones de nucletidos. Por ejemplo, el cromosoma humano ms largo,
el cromosoma nmero 1, tiene aproximadamente 220 millones de pares de bases.
21
En los organismos vivos, el ADN no suele existir como una molcula individual, sino como una pareja de
molculas estrechamente asociadas. Las dos cadenas de ADN se enroscan sobre s mismas formando una
especie de escalera de caracol, denominada doble hlice. El modelo de estructura en doble hlice fue propuesto
en 1953 por James Watson y Francis Crick (el artculo Molecular Structure of Nucleic Acids: A Structure for
Deoxyribose Nucleic Acid fue publicado el 25 de abril de 1953 en Nature), despus de obtener una imagen de la
estructura de doble hlice gracias a la refraccin por rayos X hecha por Rosalind Franklin.
22
El xito de este
modelo radicaba en su consistencia con las propiedades fsicas y qumicas del ADN. El estudio mostraba adems
que la complementariedad de bases poda ser relevante en su replicacin, y tambin la importancia de la
secuencia de bases como portadora de informacin gentica.
23
24
25
Cada unidad que se repite, el nucletido,
contiene un segmento de la estructura de soporte (azcar + fosfato), que mantiene la cadena unida, y una base,
que interacciona con la otra cadena de ADN en la hlice. En general, una base ligada a un azcar se
denomina nuclesido y una base ligada a un azcar y a uno o ms grupos fosfatos recibe el nombre
de nucletido.
Cuando muchos nucletidos se encuentran unidos, como ocurre en el ADN, el polmero resultante se
denominapolinucletido.
26
Componentes
Estructura de soporte: La estructura de soporte de una hebra de ADN est formada por unidades alternas de
grupos fosfato y azcar (desoxirribosa).
27
El azcar en el ADN es una pentosa, concretamente, la desoxirribosa.
cido fosfrico:
Enlace fosfodister. El grupo fosfato(PO
4
3-
) une el carbono 5' del azcar de unnuclesido con el carbono 3' del
siguiente.
Su frmula qumica es H
3
PO
4
. Cada nucletido puede contener uno (monofosfato: AMP), dos (difosfato: ADP) o
tres (trifosfato:ATP) grupos de cido fosfrico, aunque como monmeros constituyentes de los cidos
nucleicos slo aparecen en forma de nuclesidos monofosfato.
Desoxirribosa:
Es un monosacrido de 5 tomos de carbono (una pentosa) derivado de la ribosa, que forma parte de la
estructura de nucletidos del ADN. Su frmula es C
5
H
10
O
4
. Una de las principales diferencias entre el ADN y
el ARN es el azcar, pues en el ARN la 2-desoxirribosa del ADN es reemplazada por una pentosa alternativa,
la ribosa.
25
Las molculas de azcar se unen entre s a travs de grupos fosfato, que forman enlaces fosfodister entre los
tomos de carbono tercero (3, tres prima) y quinto (5, cinco prima) de dos anillos adyacentes de azcar. La
formacin de enlacesasimtricos implica que cada hebra de ADN tiene una direccin. En una doble hlice, la
direccin de los nucletidos en una hebra (3 5) es opuesta a la direccin en la otra hebra (5 3). Esta
organizacin de las hebras de ADN se denomina antiparalela; son cadenas paralelas, pero con direcciones
opuestas. De la misma manera, los extremos asimtricos de las hebras de ADN se denominan extremo 5 (cinco
prima) y extremo 3 (tres prima), respectivamente.
Bases nitrogenadas:
Las cuatro bases nitrogenadas mayoritarias que se encuentran en el ADN son la adenina (A), la citosina (C),
la guanina (G) y latimina (T). Cada una de estas cuatro bases est unida al armazn de azcar-fosfato a travs del
azcar para formar el nucletido completo (base-azcar-fosfato). Las bases son compuestos
heterocclicos y aromticos con dos o ms tomos de nitrgeno, y, dentro de las bases mayoritarias, se clasifican
en dos grupos: las bases pricas o purinas (adenina y guanina), derivadas de lapurina y formadas por dos anillos
unidos entre s, y las bases pirimidnicas o bases pirimdicas o pirimidinas (citosina y timina), derivadas de la
pirimidina y con un solo anillo.
25
En los cidos nucleicos existe una quinta base pirimidnica,
denominada uracilo(U), que normalmente ocupa el lugar de la timina en el ARN y difiere de sta en que carece
de un grupo metilo en su anillo. El uracilo no se encuentra habitualmente en el ADN, slo aparece raramente
como un producto residual de la degradacin de la citosina por procesos de desaminacin oxidativa.
Timina: 2, 4-dioxo, 5-metilpirimidina.
Timina:
En el cdigo gentico se representa con la letra T. Es un derivado pirimidnico con un grupo oxo en las
posiciones 2 y 4, y un grupo metil en la posicin 5. Forma el nuclesido timidina (siempre desoxitimidina, ya que
slo aparece en el ADN) y el nucletido timidilato o timidina monofosfato (dTMP). En el ADN, la timina siempre
se empareja con la adenina de la cadena complementaria mediante 2 puentes de hidrgeno, T=A. Su frmula
qumica es C
5
H
6
N
2
O
2
y su nomenclatura 2, 4-dioxo, 5-metilpirimidina.
Citosina: 2-oxo, 4-aminopirimidina.
Citosina:
En el cdigo gentico se representa con la letra C. Es un derivado pirimidnico, con un grupo amino en posicin 4
y un grupo oxo en posicin 2. Forma el nuclesido citidina (desoxicitidina en el ADN) y el nucletido citidilato o
(desoxi)citidina monofosfato (dCMP en el ADN, CMP en el ARN). La citosina siempre se empareja en el ADN con
la guanina de la cadena complementaria mediante un triple enlace, CG. Su frmula qumica es C
4
H
5
N
3
O y su
nomenclatura 2-oxo, 4 aminopirimidina. Su masa molecular es de 111,10 unidades de masa atmica. La citosina
se descubri en 1894, al aislarla del tejido del timo de carnero.
Adenina: 6-aminopurina.
Adenina:
En el cdigo gentico se representa con la letra A. Es un derivado de la purina con un grupo amino en la posicin
6. Forma el nuclesido adenosina (desoxiadenosina en el ADN) y el nucletido adenilato o (desoxi)adenosina
monofosfato (dAMP, AMP). En el ADN siempre se empareja con la timina de la cadena complementaria
mediante 2 puentes de hidrgeno, A=T. Su frmula qumica es C
5
H
5
N
5
y su nomenclatura 6-aminopurina. La
adenina, junto con la timina, fue descubierta en 1885 por el mdico alemn Albrecht Kossel.
Guanina: 6-oxo, 2-aminopurina.
Guanina:
En el cdigo gentico se representa con la letra G. Es un derivado prico con un grupo oxo en la posicin 6 y un
grupo amino en la posicin 2. Forma el nuclesido (desoxi)guanosina y el nucletido guanilato o
(desoxi)guanosina monofosfato (dGMP, GMP). La guanina siempre se empareja en el ADN con la citosina de la
cadena complementaria mediante tres enlaces de hidrgeno, GC. Su frmula qumica es C
5
H
5
N
5
O y su
nomenclatura 6-oxo, 2-aminopurina.
Tambin existen otras bases nitrogenadas (las llamadas bases nitrogenadas minoritarias), derivadas de forma
natural o sinttica de alguna otra base mayoritaria. Lo son por ejemplo la hipoxantina, relativamente abundante
en el tRNA, o la cafena, ambas derivadas de la adenina; otras, como el aciclovir, derivadas de la guanina, son
anlogos sintticos usados en terapia antiviral; otras, como una de las derivadas del uracilo, son antitumorales.
Las bases nitrogenadas tienen una serie de caractersticas que les confieren unas propiedades determinadas.
Una caracterstica importante es su carcter aromtico, consecuencia de la presencia en el anillo de dobles
enlaces en posicin conjugada. Ello les confiere la capacidad de absorber luz en la
zona ultravioleta del espectro en torno a los 260 nm, lo cual puede aprovecharse para determinar el coeficiente
de extincin del ADN y hallar la concentracin existente de los cidos nucleicos. Otra de sus caractersticas es
que presentan tautomera o isomera de grupos funcionales, debido a que un tomo de hidrgeno unido a otro
tomo puede migrar a una posicin vecina; en las bases nitrogenadas se dan dos tipos de tautomeras:
tautomera lactama-lactima, donde el hidrgeno migra del nitrgeno al oxgeno del grupo oxo (forma lactama) y
viceversa (forma lactima), y tautomera imina-amina primaria, donde el hidrgeno puede estar formando el
grupo amina (forma amina primaria) o migrar al nitrgeno adyacente (forma imina). La adenina slo puede
presentar tautomera amina-imina, la timina y el uracilo muestran tautomera doble lactama-lactima, y la
guanina y citosina pueden presentar ambas. Por otro lado, y aunque se trate de molculas apolares, las bases
nitrogenadas presentan suficiente carcter polar como para establecer puentes de hidrgeno, ya que tienen
tomos muy electronegativos (nitrgeno y oxgeno) que presentan carga parcial negativa, y tomos de
hidrgeno con carga parcial positiva, de manera que se forman dipolos que permiten que se formen
estos enlaces dbiles.
Se estima que el genoma humano haploide tiene alrededor de 3.000 millones de pares de bases. Para indicar el
tamao de las molculas de ADN se indica el nmero de pares de bases, y como derivados hay dos unidades de
medida muy utilizadas, la kilobase (kb), que equivale a 1.000 pares de bases, y la megabase (Mb), que equivale a
un milln de pares de bases.
Apareamiento de bases
Vase tambin: Par de bases
Un par de bases CG con trespuentes de hidrgeno.
Un par A=T con dos puentes de hidrgeno. Los puentes de hidrgeno se muestran como lneas discontinuas.
La dble hlice de ADN se mantiene estable mediante la formacin de puentes de hidrgeno entre las bases
asociadas a cada una de las dos hebras. Para la formacin de un enlace de hidrgeno una de las bases debe
presentar un "donador" de hidrgenos con un tomo de hidrgeno con carga parcial positiva (-NH
2
o -NH) y la
otra base debe presentar un grupo "aceptor" de hidrgenos con un tomo cargadoelectronegativamente (C=O o
N). Los puentes de hidrgeno son uniones ms dbiles que los tpicos enlaces qumicos covalentes, como los que
conectan los tomos en cada hebra de ADN, pero ms fuertes que interacciones hidrfobas individuales, enlaces
de Van der Waals, etc. Como los puentes de hidrgeno no son enlaces covalentes, pueden romperse y formarse
de nuevo de forma relativamente sencilla. Por esta razn, las dos hebras de la doble hlice pueden separarse
como una cremallera, bien por fuerza mecnica o por altatemperatura.
28
La doble hlice se estabiliza adems
por el efecto hidrofbico y el apilamiento, que no se ven influidos por la secuencia de bases del ADN.
29
Cada tipo de base en una hebra forma un enlace nicamente con un tipo de base en la otra hebra, lo que se
denominacomplementariedad de las bases. As, las purinas forman enlaces con las pirimidinas, de forma que A
se enlaza slo con T, y C slo con G. La organizacin de dos nucletidos apareados a lo largo de la doble hlice se
denomina apareamiento de bases. Este emparejamiento corresponde a la observacin ya realizada por Erwin
Chargaff (1905-2002),
30
que mostr que la cantidad de adenina era muy similar a la cantidad de timina, y que la
cantidad de citosina era igual a la cantidad de guanina en el ADN. Como resultado de esta complementariedad,
toda la informacin contenida en la secuencia de doble hebra de la hlice de ADN est duplicada en cada hebra,
lo cual es fundamental durante el proceso de replicacin del ADN. En efecto, esta interaccin reversible y
especfica entre pares de bases complementarias es crtica para todas las funciones del ADN en los organismos
vivos.
18
Como se ha indicado anteriormente, los dos tipos de pares de bases forman un nmero diferente de enlaces de
hidrgeno: A=T forman dos puentes de hidrgeno, y CG forman tres puentes de hidrgeno (ver imgenes). El
par de bases GC es por tanto ms fuerte que el par de bases AT. Como consecuencia, tanto el porcentaje de
pares de bases GC como la longitud total de la doble hlice de ADN determinan la fuerza de la asociacin entre
las dos hebras de ADN. Las dobles hlices largas de ADN con alto contenido en GC tienen hebras que
interaccionan ms fuertemente que las dobles hlices cortas con alto contenido en AT.
31
Por esta razn, las
zonas de la doble hlice de ADN que necesitan separarse fcilmente tienden a tener un alto contenido en AT,
como por ejemplo la secuencia TATAAT de la caja de Pribnow de algunos promotores.
32
En el laboratorio, la
fuerza de esta interaccin puede medirse buscando la temperatura requerida para romper los puentes de
hidrgeno, la temperatura de fusin (tambin denominado valor T
m
, del ingls melting temperature). Cuando
todas las pares de bases en una doble hlice se funden, las hebras se separan en solucin en dos hebras
completamente independientes. Estas molculas de ADN de hebra simple no tienen una nica forma comn,
sino que algunas conformaciones son ms estables que otras.
33
Otros tipos de pares de bases
Par de bases A=T de tipo Watson-Crick. En azul el donador de hidrgenos y en rojo el aceptor.
Par de bases A=T de tipo Watson-Crick reverso. En azul el donador de hidrgenos y en rojo el aceptor. Ntese
que la pirimidina ha sufrido un giro de 180 sobre el eje del carbono 6.
Existen diferentes tipos de pares de bases que se pueden formar segn el modo como se forman los puentes de
hidrgeno. Los que se observan en la doble hlice de ADN son los llamados pares de bases Watson-Crick, pero
tambin existen otros posibles pares de bases, como los denominados Hoogsteen y Wobble u oscilante, que
pueden aparecer en circunstancias particulares. Adems, para cada tipo existe a su vez el mismo par reverso, es
decir, el que se da si se gira la base pirimidnica 180 sobre su eje.
Watson-Crick (pares de bases de la doble hlice): los grupos de la base prica que intervienen
en el enlace de hidrgeno son los que corresponden a las posiciones 1 y 6 (N aceptor y -
NH
2
donador si la purina es una A) y los grupos de la base pirimidnica, los que se encuentran
en las posiciones 3 y 4 (-NH donador y C=O aceptor si la pirimidina es una T). En el par de bases
Watson-Crick reverso participaran los grupos de las posiciones 2 y 3 de la base pirimidnica
(ver imgenes).
Hoogsteen: en este caso cambian los grupos de la base prica, que ofrece una cara diferente
(posiciones 6 y 7) y que forman enlaces con los grupos de las pirimidinas de las posiciones 3 y 4
(como en Watson-Crick). Tambin puede haber Hoogsteen reversos. Con este tipo de enlace
pueden unirse A=U (Hoogsteen y Hoogsteen reverso) y A=C (Hoogsteen reverso).
Wobble u oscilante: este tipo de enlace permite que se unan guanina y citosina con un doble
enlace (G=T). La base prica (G) forma enlace con los grupos de las posiciones 1 y 6 (como en
Watson-Crick) y la pirimidina (T) con los grupos de las posiciones 2 y 3. Este tipo de enlace no
funcionara con A=C, ya que quedaran enfrentados los 2 aceptores y los 2 donadores, y slo se
podra dar en el caso inverso. Encontramos pares de bases de tipo oscilante en el ARN, durante
el apareamiento de codn y anticodn. Con este tipo de enlace pueden unirse G=U (oscilante y
oscilante reverso) y A=C (oscilante reverso).
En total, en su forma tautomrica mayoritaria, existen 28 posibles pares de bases nitrogenadas: 10 posibles
pares de bases purina-pirimidina (2 pares Watson-Crick y 2 Watson Crick reverso, 1 par Hoogsteen y 2 pares
Hoogsteen reverso, 1 par oscilante y 2 pares oscilante reverso), 7 pares homo purina-purina (A=A, G=G), 4 pares
A=G y 7 pares pirimidina-pirimidina. Esto sin contar con los pares de bases que pueden formarse si tambin
tenemos en cuenta las otras formas tautomricas minoritarias de las bases nitrogenadas; stos, adems,
pueden ser responsables de mutaciones puntuales por sustitucin de tipo transicin.
Estructura
El ADN es una molcula bicatenaria, es decir, est formada por dos cadenas dispuestas de forma antiparalela y
con las bases nitrogenadas enfrentadas. En su estructura tridimensional, se distinguen distintos niveles:
34
35
1. Estructura primaria:
Secuencia de nucletidos encadenados. Es en estas cadenas donde se encuentra la
informacin gentica, y dado que el esqueleto es el mismo para todos, la diferencia de
la informacin radica en la distinta secuencia de bases nitrogenadas. Esta secuencia
presenta un cdigo, que determina una informacin u otra, segn el orden de las
bases.
2. Estructura secundaria:
Es una estructura en doble hlice. Permite explicar el almacenamiento de la
informacin gentica y el mecanismo de duplicacin del ADN. Fue postulada por
Watson y Crick, basndose en la difraccin de rayos X que haban realizado Franklin y
Wilkins, y en la equivalencia de bases de Chargaff, segn la cual la suma de adeninas
ms guaninas es igual a la suma de timinas ms citosinas.
Es una cadena doble, dextrgira o levgira, segn el tipo de ADN. Ambas cadenas son
complementarias, pues la adenina y la guanina de una cadena se unen,
respectivamente, a la timina y la citosina de la otra. Ambas cadenas son antiparalelas,
pues el extremo 3 de una se enfrenta al extremo 5 de la homloga.
Existen tres modelos de ADN. El ADN de tipo B es el ms abundante y es el que tiene la
estructura descrita por Watson y Crick.
3. Estructura terciaria:
Se refiere a cmo se almacena el ADN en un espacio reducido, para formar
los cromosomas. Vara segn se trate de organismos procariotas o eucariotas:
2. En procariotas el ADN se pliega como una sper-hlice, generalmente en forma circular y
asociada a una pequea cantidad de protenas. Lo mismo ocurre enorgnulos celulares como
las mitocondrias y en los cloroplastos.
3. En eucariotas, dado que la cantidad de ADN de cada cromosoma es muy grande, el
empaquetamiento ha de ser ms complejo y compacto; para ello se necesita la presencia de protenas,
como las histonas y otras protenas de naturaleza no histnica (en los espermatozoides estas protenas
son las protaminas).
34
4. Estructura cuaternaria:
La cromatina presente en el ncleo tiene un grosor de 300 , pues la fibra de cromatina de 100
se enrolla formando una fibra de cromatina de 300 . El enrollamiento de
los nucleosomas recibe el nombre de solenoide. Dichos solenoides se enrollan formando la
cromatina del ncleo interfsico de la clula eucariota. Cuando la clula entra en divisin, el
ADN se compacta ms, formando as los cromosomas.
Estructuras en doble hlice
De izquierda a derecha, las estructuras de ADN A, B y Z.
El ADN existe en muchas conformaciones.
27
Sin embargo, en organismos vivos slo se han observado las
conformaciones ADN-A, ADN-By ADN-Z. La conformacin que adopta el ADN depende de su secuencia, la
cantidad y direccin de superenrollamiento que presenta, la presencia de modificaciones qumicas en las bases
y las condiciones de la solucin, tales como la concentracin de iones de metales ypoliaminas.
36
De las tres
conformaciones, la forma "B" es la ms comn en las condiciones existentes en las clulas.
37
Las dos dobles
hlices alternativas del ADN difieren en su geometra y dimensiones.
La forma "A" es una espiral que gira hacia la derecha, ms amplia que la "B", con una hendidura menor
superficial y ms amplia, y una hendidura mayor ms estrecha y profunda. La forma "A" ocurre en condiciones
no fisiolgicas en formas deshidratadas de ADN, mientras que en la clula puede producirse en apareamientos
hbridos de hebras ADN-ARN, adems de en complejos enzima-ADN.
38
39
Los segmentos de ADN en los que las bases han sido modificadas por metilacin pueden sufrir cambios
conformacionales mayores y adoptar la forma "Z". En este caso, las hebras giran alrededor del eje de la hlice en
una espiral que gira a mano izquierda, lo opuesto a la forma "B" ms frecuente.
40
Estas estructuras poco
frecuentes pueden ser reconocidas por protenas especficas que se unen a ADN-Z y posiblemente estn
implicadas en la regulacin de la transcripcin.
41
You might also like
- Problemas de intereses y descuentosDocument7 pagesProblemas de intereses y descuentosGuillermo GalvánNo ratings yet
- Derechos de Los ContribuyentesDocument5 pagesDerechos de Los ContribuyentesMirta Anayeli Ramos LopezNo ratings yet
- Act 1 2014Document2 pagesAct 1 2014Guillermo GalvánNo ratings yet
- Lista de Verificación de CFF 2016Document1 pageLista de Verificación de CFF 2016Guillermo GalvánNo ratings yet
- Idea Universo Siglos XviDocument23 pagesIdea Universo Siglos XviGuillermo GalvánNo ratings yet
- Amalgamación en Frío o Beneficio de PatioDocument2 pagesAmalgamación en Frío o Beneficio de PatioGuillermo GalvánNo ratings yet
- Volantes MamaDocument1 pageVolantes MamaGuillermo GalvánNo ratings yet
- Guía de Actividad y Rubrica de Evaluación - Tarea 2 - Genetica y BiotecnologiaDocument15 pagesGuía de Actividad y Rubrica de Evaluación - Tarea 2 - Genetica y BiotecnologiaLucia JaimesNo ratings yet
- Investigacion Medico de Planta - ResidenciaDocument150 pagesInvestigacion Medico de Planta - ResidenciaMirian BistoccoNo ratings yet
- GUÍA No.5 GENÉTICA MOLECULAR - ARN 2022Document8 pagesGUÍA No.5 GENÉTICA MOLECULAR - ARN 2022nadir steven jimenez rodriguezNo ratings yet
- Preguntas ContestadasDocument4 pagesPreguntas Contestadasnirvanita500050% (6)
- Síntesis ADNDocument7 pagesSíntesis ADNSantiago RochaNo ratings yet
- El - Cuaderno - 3 1Document9 pagesEl - Cuaderno - 3 1Nancy Pamela LoggiaNo ratings yet
- 15-16 T 16 Genética Molecular PDFDocument3 pages15-16 T 16 Genética Molecular PDFRicardo Lloyd JonesNo ratings yet
- Código GenéticoDocument6 pagesCódigo GenéticoVanessa Vera ReyesNo ratings yet
- Agraria Repaso 2019Document15 pagesAgraria Repaso 2019Alexander BényerackNo ratings yet
- Características Del Código GenéticoDocument2 pagesCaracterísticas Del Código GenéticoLenyBelénLoncomilla63% (16)
- Biología grado noveno: procesos celulares claveDocument3 pagesBiología grado noveno: procesos celulares claveLaura Sofia Salamanca JimenezNo ratings yet
- Repaso Expe 1 PARCIALDocument76 pagesRepaso Expe 1 PARCIALTefiCairellaNo ratings yet
- Introduccion A La Genetica ClinicaDocument28 pagesIntroduccion A La Genetica ClinicaGaby MoralesNo ratings yet
- Glosario de GeneticaDocument20 pagesGlosario de GeneticaE ArtunduagaNo ratings yet
- Guia 3 Biología 9° Aminoacidos y ProteinasDocument10 pagesGuia 3 Biología 9° Aminoacidos y Proteinaselizabeth ParraNo ratings yet
- Trabajo Genetica MolecularDocument10 pagesTrabajo Genetica MoleculartatianaNo ratings yet
- ADN y Genética: Docente: Laura Camila Vargas RamírezDocument23 pagesADN y Genética: Docente: Laura Camila Vargas RamírezsaritaNo ratings yet
- Genética Agrícola: Principios GenéticosDocument14 pagesGenética Agrícola: Principios GenéticosPaola Sosa WilliamsNo ratings yet
- Quiz GeneticaDocument8 pagesQuiz GeneticaWilliam Fernando Esquiivel DiazNo ratings yet
- Codificacion Del ADNDocument15 pagesCodificacion Del ADNMilkobich 05No ratings yet
- Cuadro SinopticoDocument1 pageCuadro SinopticoYexalen Pereda EstradaNo ratings yet
- CLASE 14-Transcripción y TraducciónDocument50 pagesCLASE 14-Transcripción y TraducciónAyeisa LopezNo ratings yet
- Genética 4ºesoDocument33 pagesGenética 4ºesoSanti100% (5)
- Codones de TerminoDocument79 pagesCodones de TerminoYazmin OsorioNo ratings yet
- ADN y Codigo Genetico 120929Document74 pagesADN y Codigo Genetico 120929Astrid CárdenasNo ratings yet
- Ciencias NaturalesDocument6 pagesCiencias NaturalesNarvaez Nair EliasNo ratings yet
- Ejercicios de GeneticaDocument39 pagesEjercicios de GeneticaNahuel Di RicoNo ratings yet
- Monografia-Genetica MICROBIANADocument20 pagesMonografia-Genetica MICROBIANAJc CastroNo ratings yet
- Resumen Certamen III - BioquímicaDocument17 pagesResumen Certamen III - BioquímicaFrancisco JavierNo ratings yet
- Taller de Flujo de Información Genético (Plas)Document7 pagesTaller de Flujo de Información Genético (Plas)Sebastian Orjuela ForeroNo ratings yet