You might also like
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5794)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (399)
- Partee, B. H. 2009, The Puzzle of Privative Adjectives PDFDocument13 pagesPartee, B. H. 2009, The Puzzle of Privative Adjectives PDFkhrinizNo ratings yet
- Partee, B. H. 2010, Privative Adjectives Subjsective Plus CoercionDocument13 pagesPartee, B. H. 2010, Privative Adjectives Subjsective Plus CoercionkhrinizNo ratings yet
- Are There Levels of Consciousness?: OpinionDocument9 pagesAre There Levels of Consciousness?: OpinionkhrinizNo ratings yet
- Seager, W. 1991, The Worm in The Cheese Leibniz, Consciousness and MatterDocument13 pagesSeager, W. 1991, The Worm in The Cheese Leibniz, Consciousness and MatterkhrinizNo ratings yet
- Ule, A. 2005, How Can One Catch A Thought-BirdDocument16 pagesUle, A. 2005, How Can One Catch A Thought-BirdkhrinizNo ratings yet
- Eye and Mind: Understanding the Fundamental of PaintingDocument28 pagesEye and Mind: Understanding the Fundamental of Paintingkhriniz80% (5)
- Schwitzgebel, E. 2015, If Materialism Is True, The US Is Probably ConsciousDocument25 pagesSchwitzgebel, E. 2015, If Materialism Is True, The US Is Probably ConsciouskhrinizNo ratings yet
- Twardowski, K. 1894, Zur Lehre Vom Inhalt Und GegenstandDocument123 pagesTwardowski, K. 1894, Zur Lehre Vom Inhalt Und Gegenstandkhriniz100% (1)
- Hamilton, J. F. 1972, The Photographic GrainDocument9 pagesHamilton, J. F. 1972, The Photographic GrainkhrinizNo ratings yet
- Kohler, A. 2003, Wittgenstein Meets Neuroscience (Review of Bennett & Hacker 2003)Document2 pagesKohler, A. 2003, Wittgenstein Meets Neuroscience (Review of Bennett & Hacker 2003)khrinizNo ratings yet
- Foster, J. 1993, Dennett's Rejection of DualismDocument15 pagesFoster, J. 1993, Dennett's Rejection of DualismkhrinizNo ratings yet
- Hickerson, R. 2004, An Indirect Defense of Direct RealismDocument7 pagesHickerson, R. 2004, An Indirect Defense of Direct RealismkhrinizNo ratings yet
- Capili, A. 2013, How Do We See' That Which Is Invisible'Document18 pagesCapili, A. 2013, How Do We See' That Which Is Invisible'khrinizNo ratings yet
- Praxis Vol 3, No. 2, Interview With Vincent DescombesDocument16 pagesPraxis Vol 3, No. 2, Interview With Vincent DescombeskhrinizNo ratings yet
- Slaby, J. 2007, Affective Intentionality and The Feeling BodyDocument16 pagesSlaby, J. 2007, Affective Intentionality and The Feeling BodykhrinizNo ratings yet
- Willard, D. 1994, The Integrity of The Mental Act Husserlian ReflectionsDocument12 pagesWillard, D. 1994, The Integrity of The Mental Act Husserlian ReflectionskhrinizNo ratings yet
- Mohrloff, U. 2009, Review of U. Meixner, The Two Sides of Being (Part II)Document17 pagesMohrloff, U. 2009, Review of U. Meixner, The Two Sides of Being (Part II)khrinizNo ratings yet
- Willard, D. 2011, Realism Sustained Interpreting Husserl's Progression Into IdealismDocument6 pagesWillard, D. 2011, Realism Sustained Interpreting Husserl's Progression Into IdealismkhrinizNo ratings yet
- Mohrloff, U. 2009, Review of U. Meixner, The Two Sides of Being (Part I)Document14 pagesMohrloff, U. 2009, Review of U. Meixner, The Two Sides of Being (Part I)khrinizNo ratings yet
- Parnia, S. Et Al. 2014, AWARE-AWAREness During REsuscitation A Prospective StudyDocument7 pagesParnia, S. Et Al. 2014, AWARE-AWAREness During REsuscitation A Prospective StudykhrinizNo ratings yet
- Husserl, E. (1924) 1974, Kant and The Idea of Transcendental PhilosophyDocument49 pagesHusserl, E. (1924) 1974, Kant and The Idea of Transcendental PhilosophykhrinizNo ratings yet
- Derrida - AporiasDocument49 pagesDerrida - AporiasSarah Cheng100% (1)
- Maddy, P. 2003, Second Philosophy (Lakatos Award Lecture)Document34 pagesMaddy, P. 2003, Second Philosophy (Lakatos Award Lecture)khrinizNo ratings yet
- Waller, R. R. 2012, Beyond Button Presses The Neuroscience of Free ActionsDocument23 pagesWaller, R. R. 2012, Beyond Button Presses The Neuroscience of Free ActionskhrinizNo ratings yet
- The Pharmakos Phenomenon: A Multifaceted SymbolDocument339 pagesThe Pharmakos Phenomenon: A Multifaceted SymbolkhrinizNo ratings yet
- Meixner, U. 2008, New Perspectives For A Dualistic Conception of Mental CausationDocument22 pagesMeixner, U. 2008, New Perspectives For A Dualistic Conception of Mental CausationkhrinizNo ratings yet
- Mele, A. 2012, Another Scientific Threat To Free Will?Document20 pagesMele, A. 2012, Another Scientific Threat To Free Will?khrinizNo ratings yet
- Sellars, W. 1956, Empiricism & The Philosophy of MindDocument39 pagesSellars, W. 1956, Empiricism & The Philosophy of MindkhrinizNo ratings yet
- Soulez, A. 1989, Wittgenstein & Phenomenology Or: Two Languages For One WittgensteinDocument27 pagesSoulez, A. 1989, Wittgenstein & Phenomenology Or: Two Languages For One WittgensteinkhrinizNo ratings yet
- Zahavi, D. 2014, Empathy & Other-Directed IntentionalityDocument14 pagesZahavi, D. 2014, Empathy & Other-Directed Intentionalitykhriniz100% (1)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (894)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (587)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (265)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (73)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (344)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2219)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (119)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- Growth PredictionDocument101 pagesGrowth PredictionKristty Magallanes100% (1)
- Chapter 1Document6 pagesChapter 1Hema Priya AdityanNo ratings yet
- Dental instruments for extracting teethDocument18 pagesDental instruments for extracting teethKateryna BondarenkoNo ratings yet
- Miss Evers Boys Draft 4Document2 pagesMiss Evers Boys Draft 4api-291172102No ratings yet
- Ergogenic Aids 1Document24 pagesErgogenic Aids 1unattractiveyouNo ratings yet
- Teva PharmaceuticalDocument17 pagesTeva PharmaceuticalGanesh VedhachalamNo ratings yet
- Retainer types and uses in orthodonticsDocument6 pagesRetainer types and uses in orthodonticsSahana RangarajanNo ratings yet
- Guide Sondes Logiq s8Document4 pagesGuide Sondes Logiq s8mahfoudNo ratings yet
- Cardiac CycleDocument2 pagesCardiac CycleU Than HanNo ratings yet
- 1 Wellness For EyesDocument5 pages1 Wellness For Eyesanna7290100% (4)
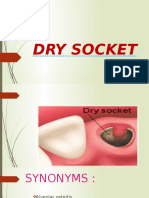
- Dry SocketDocument20 pagesDry SocketHosuru SwethaNo ratings yet
- As 5013.2-2007 Food Microbiology Microbiology of Food and Animal Feeding Stuffs - Horizontal Method For The eDocument10 pagesAs 5013.2-2007 Food Microbiology Microbiology of Food and Animal Feeding Stuffs - Horizontal Method For The eSAI Global - APAC100% (1)
- MRCP UK PACES EXAM GUIDEDocument124 pagesMRCP UK PACES EXAM GUIDEJayachandran P KNo ratings yet
- Toxicology Procedures ManualDocument227 pagesToxicology Procedures ManualBenjel AndayaNo ratings yet
- MidazolamDocument18 pagesMidazolamHarnugrahanto AankNo ratings yet
- Autologous Transfusion Strategies to Reduce Allogeneic Blood UseDocument31 pagesAutologous Transfusion Strategies to Reduce Allogeneic Blood Usethalida24100% (1)
- Statement On Controlled Organ Donation After Circulatory DeathDocument10 pagesStatement On Controlled Organ Donation After Circulatory DeathHeidi ReyesNo ratings yet
- Compartment SyndromeDocument29 pagesCompartment SyndromeFazmial UjirNo ratings yet
- Hypertension: Rojina Bhurtel Lecturer MmihsDocument36 pagesHypertension: Rojina Bhurtel Lecturer MmihsRojina Bhurtel100% (2)
- Obat SpesialisDocument16 pagesObat SpesialisLAILATUL AFIYAHNo ratings yet
- Beximco Pharma FinalDocument14 pagesBeximco Pharma FinalAli Asgor Raton67% (3)
- Best Practice & Research Clinical Obstetrics and GynaecologyDocument12 pagesBest Practice & Research Clinical Obstetrics and GynaecologyGrit WingsNo ratings yet
- Adr FormDocument2 pagesAdr FormSachin Kumar PrajapatiNo ratings yet
- Ecg InterpretationDocument3 pagesEcg Interpretationman0billi0% (1)
- Candida Albicans and IridologyDocument8 pagesCandida Albicans and IridologyFRANCISCO JAVIER GALEANONo ratings yet
- Respiratory Distress SyndromeDocument54 pagesRespiratory Distress SyndromeKristine CaringalNo ratings yet
- 1 - General Indications and Contraindications - 2019 - Lumbar Interbody FusionsDocument12 pages1 - General Indications and Contraindications - 2019 - Lumbar Interbody FusionsSergiu MalinNo ratings yet
- DetailsDocument21 pagesDetailskanthavelNo ratings yet
- Morning Report (Cardiology) : On Duty: Nadya Julaifa, Nadya Julaifa Supervisor: Dr. Yusra Pintaningrum, Sp. JP (K) FIHADocument16 pagesMorning Report (Cardiology) : On Duty: Nadya Julaifa, Nadya Julaifa Supervisor: Dr. Yusra Pintaningrum, Sp. JP (K) FIHAAdisti NurutamiNo ratings yet