You might also like
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5794)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1090)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (890)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (587)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (344)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (119)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (399)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2219)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (265)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (73)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- 02Q PDFDocument1 page02Q PDFFCiênciasNo ratings yet
- Lambers, Oliveira 2019 Plant Physiological Ecology 3rdDocument755 pagesLambers, Oliveira 2019 Plant Physiological Ecology 3rdLuisFrancisco BustamanteNo ratings yet
- 11 - Plant Defenses Against Biotic Stresses PDFDocument57 pages11 - Plant Defenses Against Biotic Stresses PDFFCiênciasNo ratings yet
- 04Q PDFDocument2 pages04Q PDFFCiênciasNo ratings yet
- Amber 15Document883 pagesAmber 15FCiênciasNo ratings yet
- 9 - Responses Adaptations To Abiotic Stresses PDFDocument62 pages9 - Responses Adaptations To Abiotic Stresses PDFFCiênciasNo ratings yet
- 07Q PDFDocument1 page07Q PDFFCiênciasNo ratings yet
- 4 - Phytohormones and Elicitor Molecules - ABSCISIC ACID PDFDocument20 pages4 - Phytohormones and Elicitor Molecules - ABSCISIC ACID PDFFCiênciasNo ratings yet
- 6 - Mineral Nutrition and Solute Transport PDFDocument19 pages6 - Mineral Nutrition and Solute Transport PDFFCiênciasNo ratings yet
- 10 - Signal Perception and Transduction PDFDocument52 pages10 - Signal Perception and Transduction PDFFCiênciasNo ratings yet
- 5 - Phytohormones and Elicitor Molecules - ETHYLENE BRASSINOSTEROIDS PDFDocument33 pages5 - Phytohormones and Elicitor Molecules - ETHYLENE BRASSINOSTEROIDS PDFFCiênciasNo ratings yet
- 2 - Phytohormones and Elicitor Molecules - CYTOKININS PDFDocument21 pages2 - Phytohormones and Elicitor Molecules - CYTOKININS PDFFCiênciasNo ratings yet
- 2 - Phytohormones and Elicitor Molecules - CYTOKININS PDFDocument21 pages2 - Phytohormones and Elicitor Molecules - CYTOKININS PDFFCiênciasNo ratings yet
- 06Q PDFDocument1 page06Q PDFFCiênciasNo ratings yet
- 8 - Secondary Metabolites - TERPENOIDS ALKALOIDS PHENOLICS PDFDocument82 pages8 - Secondary Metabolites - TERPENOIDS ALKALOIDS PHENOLICS PDFFCiênciasNo ratings yet
- 3 - Phytohormones and Elicitor Molecules - GIBBERELLINS PDFDocument18 pages3 - Phytohormones and Elicitor Molecules - GIBBERELLINS PDFFCiênciasNo ratings yet
- 08Q PDFDocument1 page08Q PDFFCiênciasNo ratings yet
- 01Q PDFDocument1 page01Q PDFFCiênciasNo ratings yet
- Detecção de Mutações - Genotipagem PDFDocument24 pagesDetecção de Mutações - Genotipagem PDFFCiênciasNo ratings yet
- 03Q PDFDocument2 pages03Q PDFFCiênciasNo ratings yet
- 7.02 Recombinant DNA Methods Spring 2005 Exam Study Questions Answer KeyDocument10 pages7.02 Recombinant DNA Methods Spring 2005 Exam Study Questions Answer KeyFCiênciasNo ratings yet
- Real-Time PCR Com Perguntas PDFDocument27 pagesReal-Time PCR Com Perguntas PDFFCiênciasNo ratings yet
- 05Q PDFDocument1 page05Q PDFFCiênciasNo ratings yet
- Exercícios - Techniques To Measure Changes in Gene Expression PDFDocument30 pagesExercícios - Techniques To Measure Changes in Gene Expression PDFFCiênciasNo ratings yet
- Exercício Mutagénese Sítio Dirigida - Questões PDFDocument25 pagesExercício Mutagénese Sítio Dirigida - Questões PDFFCiênciasNo ratings yet
- EXERCICIO Degenerate - Probes - Exerecicio2013 PDFDocument4 pagesEXERCICIO Degenerate - Probes - Exerecicio2013 PDFFCiênciasNo ratings yet
- EXERCICIOS Cloning PCR PRODUCTS Exercício PDFDocument9 pagesEXERCICIOS Cloning PCR PRODUCTS Exercício PDFFCiênciasNo ratings yet
- 06 MMB2013 PDFDocument23 pages06 MMB2013 PDFFCiênciasNo ratings yet
- Alcohol - Metab p450 PDFDocument17 pagesAlcohol - Metab p450 PDFFCiênciasNo ratings yet
- 1ºteste 2013 2014 Catarina Cunha PDFDocument4 pages1ºteste 2013 2014 Catarina Cunha PDFFCiênciasNo ratings yet
- Additional Biogeochemical CycleDocument9 pagesAdditional Biogeochemical CycleYngrid Bless Z. CadenaNo ratings yet
- The Role of Agro Inputs Rhizobia P and K and Critical Stages of Application For Improved Soybean Yield and BiomassDocument29 pagesThe Role of Agro Inputs Rhizobia P and K and Critical Stages of Application For Improved Soybean Yield and BiomasswindamarpaungNo ratings yet
- Igcse Biology UNIT 19.4Document38 pagesIgcse Biology UNIT 19.4Ajinkya dangeNo ratings yet
- AP Biology Bacteria and VirusesDocument51 pagesAP Biology Bacteria and VirusesMichael TsaiNo ratings yet
- Differences Between Prokaryotes and EukaryotesDocument256 pagesDifferences Between Prokaryotes and EukaryotesMuhammad Syamsul ArifinNo ratings yet
- Biogeochemical Cycles Webquest ActivityDocument5 pagesBiogeochemical Cycles Webquest Activitykate nicole labandriaNo ratings yet
- Can Top-Down Controls Expand The Ecological Niche of Marine N Fixers?Document11 pagesCan Top-Down Controls Expand The Ecological Niche of Marine N Fixers?Neha_ Sharma08No ratings yet
- Maní en El Ensamblaje Microbiano, Los Exudados de Raíces y La Absorción de Nitrógeno en El ManíDocument9 pagesManí en El Ensamblaje Microbiano, Los Exudados de Raíces y La Absorción de Nitrógeno en El ManíBruno ReyesNo ratings yet
- Rhizobium Root Nodules & Nitrogen FixationDocument4 pagesRhizobium Root Nodules & Nitrogen Fixationzoba85100% (1)
- Soil MicrobiologyDocument30 pagesSoil MicrobiologyRajan Bhatt, Ph.D (Soil Science)92% (12)
- Biofertilizer For Crop Production and Soil Fertility: August 2018Document9 pagesBiofertilizer For Crop Production and Soil Fertility: August 2018GnanakumarNo ratings yet
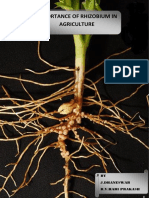
- Importance of Rhizobium in AgricultureDocument16 pagesImportance of Rhizobium in AgricultureDhaneswarNo ratings yet
- 12 Mineral NutritionDocument5 pages12 Mineral NutritionDe DasNo ratings yet
- Soil BacteriaDocument2 pagesSoil Bacteriaani4 samNo ratings yet
- PblockDocument20 pagesPblockManish kumarNo ratings yet
- Home Gardeners Guide To Soils and Fertilizers - WSUpdfDocument26 pagesHome Gardeners Guide To Soils and Fertilizers - WSUpdfDNo ratings yet
- Mineral Nutrition BioHackDocument8 pagesMineral Nutrition BioHackMohammed HamedNo ratings yet
- Biofertilizer 4th LevelDocument24 pagesBiofertilizer 4th LevelSamah RamadanNo ratings yet
- Engenvi NotesDocument12 pagesEngenvi NotesMariaNo ratings yet
- Balancing Global BiodiversityDocument11 pagesBalancing Global BiodiversityAref DahabrahNo ratings yet
- Rural Agricultural Work Experience in TripuraDocument59 pagesRural Agricultural Work Experience in TripuraSamuel Davis100% (1)
- Nitrogen CycleDocument15 pagesNitrogen CycleZoya ShaikhNo ratings yet
- Unit-1 Natural CyclesDocument13 pagesUnit-1 Natural CyclesAkshit KansalNo ratings yet
- Soil FertilityDocument84 pagesSoil FertilityakNo ratings yet
- 37: Soil and Plant NutritionDocument7 pages37: Soil and Plant NutritionyugawadNo ratings yet
- Module 5Document14 pagesModule 5Melody GarciaNo ratings yet
- Useful Microbes: Project Done By: Abdullah, Barrry, Andrew, Freda (Group 3)Document8 pagesUseful Microbes: Project Done By: Abdullah, Barrry, Andrew, Freda (Group 3)Muhammad AbdullahNo ratings yet
- Mikrobiologi Dasar-Pertemuan 2Document22 pagesMikrobiologi Dasar-Pertemuan 2Wahyu Aji SantosoNo ratings yet
- Biogeochemical Cycles For APESDocument42 pagesBiogeochemical Cycles For APESjohnosborneNo ratings yet