You might also like
- Role of Hyaluronan in Angiogenesis and Its Utility To Angiogenic TissueDocument12 pagesRole of Hyaluronan in Angiogenesis and Its Utility To Angiogenic TissueCarlos LabradaNo ratings yet
- Systems For Therapeutic Angiogenesis in Tissue EngineeringDocument9 pagesSystems For Therapeutic Angiogenesis in Tissue EngineeringCarlos LabradaNo ratings yet
- Endothelial Extracellular MatrixDocument17 pagesEndothelial Extracellular MatrixCarlos LabradaNo ratings yet
- Current Rehabilitation Strategy Clinical Evidence For Erection Recovery After Radical ProstatectomyDocument8 pagesCurrent Rehabilitation Strategy Clinical Evidence For Erection Recovery After Radical ProstatectomyCarlos LabradaNo ratings yet
- 2001 - Wnek - Arterial Smooth Muscle Cell Proliferation On A Novel Biomimicking Biodegradable Vascular Graft ScaffoldDocument12 pages2001 - Wnek - Arterial Smooth Muscle Cell Proliferation On A Novel Biomimicking Biodegradable Vascular Graft ScaffoldCarlos LabradaNo ratings yet
- Research Paper NS11021, A Novel Opener of Large-Conductance Ca - Activated K Channels, Enhances Erectile Responses in RatsDocument12 pagesResearch Paper NS11021, A Novel Opener of Large-Conductance Ca - Activated K Channels, Enhances Erectile Responses in RatsCarlos LabradaNo ratings yet
- Dmso Libro FelizDocument110 pagesDmso Libro Felizsilke000100% (3)
- The Role of Isoforms of Soluble Guanylyl Cyclase in Relaxation of Vascular Smooth Muscle Cells and Corpora Cavernosa - ThesisKellyDecaluweDocument246 pagesThe Role of Isoforms of Soluble Guanylyl Cyclase in Relaxation of Vascular Smooth Muscle Cells and Corpora Cavernosa - ThesisKellyDecaluweCarlos LabradaNo ratings yet
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5784)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (890)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (399)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (587)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (265)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (72)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (344)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2219)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (119)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- Inclusive E-Service or Risk of Digital Divide The Case of National ICT Policy 2018 of BangladeshDocument11 pagesInclusive E-Service or Risk of Digital Divide The Case of National ICT Policy 2018 of BangladeshInternational Journal of Innovative Science and Research Technology100% (1)
- Metal Oxides Semiconductor CeramicsDocument14 pagesMetal Oxides Semiconductor Ceramicsumarasad1100% (1)
- V60 Ventilator Specifications PDFDocument4 pagesV60 Ventilator Specifications PDFJonathan Issac Dominguez RamirezNo ratings yet
- About Topsøe - and What We DoDocument20 pagesAbout Topsøe - and What We DoAbhishek ChaudharyNo ratings yet
- Unit 01 Family Life Lesson 1 Getting Started - 2Document39 pagesUnit 01 Family Life Lesson 1 Getting Started - 2Minh Đức NghiêmNo ratings yet
- Balzac and the Little Chinese Seamstress: A Journey During the Cultural RevolutionDocument4 pagesBalzac and the Little Chinese Seamstress: A Journey During the Cultural RevolutionRogona 123No ratings yet
- Brochure HorticultureDocument46 pagesBrochure HorticulturezulfiqaralimalikNo ratings yet
- Optitex Com Products 2d and 3d Cad SoftwareDocument12 pagesOptitex Com Products 2d and 3d Cad SoftwareFaathir Reza AvicenaNo ratings yet
- EOD Stanags Overview April 2021Document12 pagesEOD Stanags Overview April 2021den mas paratate leo egoNo ratings yet
- Adapting Cognitive Behavioral Techniques To Address Anxiety and Depression in Cognitively Able Emerging Adults On The Autism SpectrumDocument3 pagesAdapting Cognitive Behavioral Techniques To Address Anxiety and Depression in Cognitively Able Emerging Adults On The Autism SpectrumVini PezzinNo ratings yet
- Komposit UHMWPE Sebagai Alternatif Bantalan Rel Kereta Api: Abel Evan, Alia Kristika, Farid Mulia LatiefDocument11 pagesKomposit UHMWPE Sebagai Alternatif Bantalan Rel Kereta Api: Abel Evan, Alia Kristika, Farid Mulia LatiefAlia KristikaNo ratings yet
- Explore Spanish Lesson Plan - AnimalsDocument8 pagesExplore Spanish Lesson Plan - Animalsapi-257582917No ratings yet
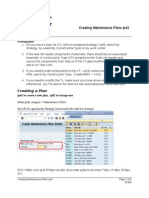
- JA Ip42 Creating Maintenance PlansDocument8 pagesJA Ip42 Creating Maintenance PlansvikasbumcaNo ratings yet
- HIS 101 Home Work 2Document10 pagesHIS 101 Home Work 2Nabil HussainNo ratings yet
- Pankaj Screener 10 Oct 2014Document127 pagesPankaj Screener 10 Oct 2014Sadul Singh Naruka100% (1)
- Component 2 Learner Statement Y2Document6 pagesComponent 2 Learner Statement Y2api-426152133No ratings yet
- Perbandingan Penggunaan Refrigeran R22 Dengan R407 Pada Performa ACDocument15 pagesPerbandingan Penggunaan Refrigeran R22 Dengan R407 Pada Performa ACYuli RahmawatiNo ratings yet
- Albert PikeDocument6 pagesAlbert Pikeapi-302575383No ratings yet
- Lorain Schools CEO Finalist Lloyd MartinDocument14 pagesLorain Schools CEO Finalist Lloyd MartinThe Morning JournalNo ratings yet
- SID-2AF User Manual English V3.04Document39 pagesSID-2AF User Manual English V3.04om_zahidNo ratings yet
- EDUC 5 - QuestionairesDocument7 pagesEDUC 5 - QuestionairesWilliam RanaraNo ratings yet
- Gabriel Nobre de Souza - Groningen Mini-MetropolisDocument9 pagesGabriel Nobre de Souza - Groningen Mini-MetropolisGabrielNobredeSouzaNo ratings yet
- Fire Pump System Test ReportDocument12 pagesFire Pump System Test Reportcoolsummer1112143100% (2)
- MARCOMDocument35 pagesMARCOMDrei SalNo ratings yet
- Effectiveness of Using High Gas Pressure in A Blast Furnace As A Means of Intensifying The Smelting OperationDocument10 pagesEffectiveness of Using High Gas Pressure in A Blast Furnace As A Means of Intensifying The Smelting Operationchandan kumar100% (1)
- CELTA Pre-Interview Grammar, Vocabulary and Pronunciation ExercisesDocument4 pagesCELTA Pre-Interview Grammar, Vocabulary and Pronunciation ExercisesMichelJorge100% (2)
- Gatk Pipeline Presentation: From Fastq Data To High Confident VariantsDocument8 pagesGatk Pipeline Presentation: From Fastq Data To High Confident VariantsSampreeth ReddyNo ratings yet
- Open MPDocument30 pagesOpen MPmacngocthanNo ratings yet
- ccpc15 Supportive and Preventive WorkbookDocument30 pagesccpc15 Supportive and Preventive WorkbookJeremy HamptonNo ratings yet
- Automated Crime Reporting SystemDocument101 pagesAutomated Crime Reporting SystemDeepak Kumar60% (10)