You might also like
- Complejo Mayor de HistocompatibilidadDocument16 pagesComplejo Mayor de Histocompatibilidadmsfts26No ratings yet
- Complejo de Mayor HistocompatibilidadDocument7 pagesComplejo de Mayor HistocompatibilidadJonatán Eliseo Morales JeronimoNo ratings yet
- Artículo Complejo Mayor de HistocompatibilidadDocument4 pagesArtículo Complejo Mayor de HistocompatibilidadRebeca Saltos CandelaNo ratings yet
- Patógenos orofaríngeos y piel: Estreptococos y EstafilococosDocument16 pagesPatógenos orofaríngeos y piel: Estreptococos y EstafilococosLiz ZacariasNo ratings yet
- Bacterias ResumenDocument74 pagesBacterias ResumenRamón H. JuárezNo ratings yet
- Lupus Eritematoso SistemicoDocument20 pagesLupus Eritematoso SistemicoTaeTJMietCZNo ratings yet
- NeurocisticercosisDocument2 pagesNeurocisticercosisMaira Vargas OdicioNo ratings yet
- Fasciola HepaticaDocument4 pagesFasciola HepaticaNoemi NaranjoNo ratings yet
- Aparato de GolgiDocument3 pagesAparato de Golgiماريا أنطونيا تابوردا ريفيرا100% (1)
- Virus de La RabiaDocument23 pagesVirus de La RabiaFabricio DilasNo ratings yet
- IMA Causas Diagnostico TratamientoDocument15 pagesIMA Causas Diagnostico TratamientoHernan RodriguezNo ratings yet
- La Hematopoyesis: Formación de células sanguíneasDocument16 pagesLa Hematopoyesis: Formación de células sanguíneasblasterplusNo ratings yet
- MiasisDocument50 pagesMiasisAmada Angel VillanuevaNo ratings yet
- Trabajo de Sistema Inmune MicrobiologiaDocument4 pagesTrabajo de Sistema Inmune MicrobiologiaCesar Ricardo Zuñiga100% (1)
- Unidad 4 Procesamiento y Presentación Del AntígenoDocument29 pagesUnidad 4 Procesamiento y Presentación Del AntígenolitorNo ratings yet
- Fisiopatología de La Tuberculosis - CHECKMEDDocument13 pagesFisiopatología de La Tuberculosis - CHECKMEDgoldenmemberNo ratings yet
- Trypanosoma SPDocument30 pagesTrypanosoma SPnikonhp100% (1)
- Dengue ExpoDocument18 pagesDengue ExpoUbaldoRuizNo ratings yet
- SEMINARIOINFLAMACIÓNDocument52 pagesSEMINARIOINFLAMACIÓNMark Vertiz CernaNo ratings yet
- LinfáticoDocument34 pagesLinfáticoFelipe Aguilera ZúñigaNo ratings yet
- Hipersensibilidad III y IV ResumenDocument16 pagesHipersensibilidad III y IV ResumenMilad JiloNo ratings yet
- Aplicaciones de La Células MadresDocument9 pagesAplicaciones de La Células MadresluisNo ratings yet
- Complejo Mayor de HistocompatibilidadDocument2 pagesComplejo Mayor de Histocompatibilidadshantal liberatoNo ratings yet
- Patogenia Del VihDocument6 pagesPatogenia Del Vihandiite100% (1)
- Inmunología del rechazo agudoDocument13 pagesInmunología del rechazo agudokeylaNo ratings yet
- Resumen CHM y TrasplanteDocument2 pagesResumen CHM y TrasplanteBenjamin F. Sandi VigabrielNo ratings yet
- Síndrome de Reye - Jose Armando Actual RamosDocument4 pagesSíndrome de Reye - Jose Armando Actual RamosArmando Actual RamosNo ratings yet
- Sindrome de GoodpastureDocument3 pagesSindrome de GoodpastureRicardo ContrerasNo ratings yet
- Respiratorio EmbrioDocument47 pagesRespiratorio EmbrioRosa Angelica Failoc PiscoyaNo ratings yet
- Infecciones OportunistasDocument64 pagesInfecciones Oportunistasjrvg2007@gmail.comNo ratings yet
- MENINGITISDocument23 pagesMENINGITISEdwind Henry Infante SanchezNo ratings yet
- Fiebre de Origen DesconocidoDocument10 pagesFiebre de Origen DesconocidoDenisse FurlanNo ratings yet
- Semiologia Dermatologica UvmDocument16 pagesSemiologia Dermatologica UvmGiannina Francesca Lagomarsino GarateNo ratings yet
- Endocarditis InfecciosaDocument7 pagesEndocarditis InfecciosaAnhuar FerrerNo ratings yet
- Cap. 55 Funciones Motoras de La Médula Espinal - Reflejos MedularesDocument10 pagesCap. 55 Funciones Motoras de La Médula Espinal - Reflejos MedularesLinda Andreita Rosales Rosales0% (1)
- p1.2 Relacion Parasito-Hospedero 08-03-2018 Dr. AguirreDocument6 pagesp1.2 Relacion Parasito-Hospedero 08-03-2018 Dr. AguirreRodrigo Ancalle ZuñigaNo ratings yet
- Cisticercosis PorcinaDocument11 pagesCisticercosis PorcinaCarmelita huancas ramirezNo ratings yet
- Cuadro Comparativo de Caracteristicas de Las Moleculas Del Complejo Mayor de HistoccompatibilidadDocument3 pagesCuadro Comparativo de Caracteristicas de Las Moleculas Del Complejo Mayor de HistoccompatibilidadGessie Licona100% (1)
- Polineuropatias Diagnostico DiferencialDocument102 pagesPolineuropatias Diagnostico DiferencialmarlingaxNo ratings yet
- MII_EpidemiologiaDocument4 pagesMII_EpidemiologiaFrancisco Riquelme UrzuaNo ratings yet
- DengueDocument23 pagesDengueBRENDA ZURITA VEIZAGA100% (1)
- Arenavirus: virus causantes de fiebres hemorrágicasDocument10 pagesArenavirus: virus causantes de fiebres hemorrágicasErik Zenteno100% (1)
- Guia de Pericarditis 2019Document23 pagesGuia de Pericarditis 2019Diego GonzalesNo ratings yet
- Ensayo Fisiopatología AteroesclerosisDocument8 pagesEnsayo Fisiopatología AteroesclerosisJavier RodríguezNo ratings yet
- Hiperinsulinismo CongenitoDocument31 pagesHiperinsulinismo CongenitoJ Francisco GarcíaNo ratings yet
- PEDICULOSISDocument4 pagesPEDICULOSISalbianny0% (1)
- Meningitis BacterianaDocument22 pagesMeningitis BacterianaRené Rubio LópezNo ratings yet
- Envoltura NuclearDocument12 pagesEnvoltura NuclearBikner Leon ParedesNo ratings yet
- Sindrome Duplo YDocument4 pagesSindrome Duplo YjersicNo ratings yet
- Expo InmunoglobulinasDocument38 pagesExpo InmunoglobulinasManuel López Cobos100% (1)
- InmunodeficienciasDocument18 pagesInmunodeficienciasDaniela ArdilaNo ratings yet
- Purpura TrombocitopenicaDocument6 pagesPurpura TrombocitopenicaOlivia MaroNo ratings yet
- Mecanismo de Patogenicidad BacterianaDocument3 pagesMecanismo de Patogenicidad BacterianaKeyla Vente VelascoNo ratings yet
- Inmunidad AdaptativaDocument6 pagesInmunidad AdaptativaRafael Rovira Gómez-SánchezNo ratings yet
- Virus Arenavirus: Características y Enfermedades ZoonóticasDocument12 pagesVirus Arenavirus: Características y Enfermedades ZoonóticasBarbara CastilloNo ratings yet
- Procesamiento Presentacion Antigenica PDFDocument28 pagesProcesamiento Presentacion Antigenica PDFRichard Bravo CéspedesNo ratings yet
- 6 Activacion de LinfocitosDocument52 pages6 Activacion de LinfocitosRocio GallegosNo ratings yet
- Pancreas PatologiasDocument115 pagesPancreas PatologiasAllonsiny GutierrezNo ratings yet
- Enfermedades Autoinmunes Sistémicas - EnfermosDocument62 pagesEnfermedades Autoinmunes Sistémicas - EnfermosNorberto Ortego Centeno100% (1)
- HISTOCOMPATIBILIDADDocument24 pagesHISTOCOMPATIBILIDADRaquel Lopez AndresNo ratings yet
- Cerveza Alianza UniversitarioDocument1 pageCerveza Alianza UniversitarioMao NgNo ratings yet
- Alianza LimaDocument3 pagesAlianza LimaMao NgNo ratings yet
- Gremio de Recolectores de Camarones de Los Ríos de La Provincia de CañeteDocument2 pagesGremio de Recolectores de Camarones de Los Ríos de La Provincia de CañeteMao NgNo ratings yet
- Introduccion Por Medicamentos - DibujosDocument5 pagesIntroduccion Por Medicamentos - DibujosMao NgNo ratings yet
- Miriam Roxsana Candela MascaroDocument36 pagesMiriam Roxsana Candela MascaroMao NgNo ratings yet
- Catálogo de productos agrícolas Tellus CorpDocument9 pagesCatálogo de productos agrícolas Tellus CorpMao NgNo ratings yet
- Doc5 Borrar BorrarDocument10 pagesDoc5 Borrar BorrarMao NgNo ratings yet
- Sumary SidaDocument10 pagesSumary SidaMao NgNo ratings yet
- Helados Vane VaneDocument15 pagesHelados Vane VaneMao Ng100% (1)
- Medidas BiométricasDocument8 pagesMedidas BiométricasMao Ng50% (2)
- Danzas Del PerúDocument8 pagesDanzas Del PerúMao NgNo ratings yet
- Lunahuana ArreglarDocument38 pagesLunahuana ArreglarMao NgNo ratings yet
- Administración de EmpresasDocument2 pagesAdministración de EmpresasMao NgNo ratings yet
- Lanza Mien ToDocument6 pagesLanza Mien ToMao NgNo ratings yet
- Gremio de Recolectores de Camarones de Los Ríos de La Provincia de CañeteDocument1 pageGremio de Recolectores de Camarones de Los Ríos de La Provincia de CañeteMao NgNo ratings yet
- Ala DeltaDocument2 pagesAla DeltaMao NgNo ratings yet
- Reglamento Nacional de Transito Borrar BorrarDocument8 pagesReglamento Nacional de Transito Borrar BorrarMao NgNo ratings yet
- Diversos TrabajosDocument3 pagesDiversos TrabajosMao NgNo ratings yet
- Albun de Animales BorrarDocument2 pagesAlbun de Animales BorrarMao NgNo ratings yet
- Administración y DerechoDocument5 pagesAdministración y DerechoMao NgNo ratings yet
- Actuación en FerarmacologíaDocument2 pagesActuación en FerarmacologíaMao NgNo ratings yet
- Adverbios de Frecuencia InglesDocument5 pagesAdverbios de Frecuencia InglesMao Ng100% (1)
- Actos Prohibidos Al Tutor - TipeoDocument4 pagesActos Prohibidos Al Tutor - TipeoMao Ng100% (1)
- Alan Garcia Perez - BorrarDocument2 pagesAlan Garcia Perez - BorrarMao NgNo ratings yet
- Actividades Recreativas Del PROMSA BorrarDocument1 pageActividades Recreativas Del PROMSA BorrarMao NgNo ratings yet
- Cultura Tiahuanaco TripticoDocument2 pagesCultura Tiahuanaco TripticoMao Ng100% (4)
- Alcaldes de ImperialDocument4 pagesAlcaldes de ImperialMao NgNo ratings yet
- Visual Studio 2010Document13 pagesVisual Studio 2010Mao NgNo ratings yet
- Programas MicrosoftDocument2 pagesProgramas MicrosoftMao NgNo ratings yet
- Articulación LingüísticaDocument1 pageArticulación LingüísticaMao NgNo ratings yet
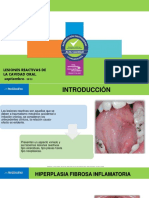
- Lesiones reactivas de la cavidad oral: Hiperplasia fibrosa inflamatoriaDocument18 pagesLesiones reactivas de la cavidad oral: Hiperplasia fibrosa inflamatoriaAllan OrtegaNo ratings yet
- Recolecion de Aceites Usados en BucaramangaDocument5 pagesRecolecion de Aceites Usados en BucaramangaFredy RuizNo ratings yet
- Trabajo en AlturaDocument45 pagesTrabajo en AlturaPaes RodrisonNo ratings yet
- Fabio Aldair MontañoDocument1 pageFabio Aldair MontañoFabio Aldair Montano MorenoNo ratings yet
- Hoope One Pager (Spanish)Document1 pageHoope One Pager (Spanish)Rubén Carlos Andrés VGNo ratings yet
- Cuadro ComparativoDocument3 pagesCuadro ComparativoKarolina Bayona Perez100% (1)
- Conceptos Basicos de FarmacologiaDocument17 pagesConceptos Basicos de FarmacologiaEric MartinezNo ratings yet
- I-SSO-005 - Procedimiento de Limpieza en PlantaDocument5 pagesI-SSO-005 - Procedimiento de Limpieza en PlantaOscar FiallosNo ratings yet
- Choquehuanca 2019 Estandarización de Elisa Anti LeishmaniaDocument14 pagesChoquehuanca 2019 Estandarización de Elisa Anti LeishmaniaBeatriz GTNo ratings yet
- Psc-304 Psicopatologia II 6 Febrero 2020 28 de Febrero 2020Document17 pagesPsc-304 Psicopatologia II 6 Febrero 2020 28 de Febrero 2020kery lantigua100% (1)
- Infografía de La Historia ClínicaDocument3 pagesInfografía de La Historia ClínicaMayra Gonzalez100% (1)
- Conciliacion BancariaDocument37 pagesConciliacion BancariaDanny PanoccatitoNo ratings yet
- Códigos de sanación numéricosDocument16 pagesCódigos de sanación numéricosReikiMaster Marcela La Serena100% (1)
- IasoTea ESDocument1 pageIasoTea ESsgranada24No ratings yet
- Plan de Reducción de Riesgos 2022Document27 pagesPlan de Reducción de Riesgos 2022Fabiola VelizNo ratings yet
- Juan Antonio Seda.Document16 pagesJuan Antonio Seda.Crys TorresNo ratings yet
- TROMBOCITOPENIADocument24 pagesTROMBOCITOPENIAAmilkar Anticona CamayocNo ratings yet
- El Servicio de Tutoria en Educacion Inicial Parte 1Document16 pagesEl Servicio de Tutoria en Educacion Inicial Parte 1Juana Elisa Dioses Rizzi75% (4)
- Actividad Evaluativa - Eje 3 SexualidadDocument8 pagesActividad Evaluativa - Eje 3 SexualidadMafe Perea50% (2)
- El Papel Que Desempeñan Los Paseadores de Perros en La Transmisión Del SARS-CoV-2 Una Revisión Desde La Salud Pública VeterinariaDocument25 pagesEl Papel Que Desempeñan Los Paseadores de Perros en La Transmisión Del SARS-CoV-2 Una Revisión Desde La Salud Pública VeterinariaVane ValenciaNo ratings yet
- Versión Reducida Del Boston Naming Test Tesis Genetica Del TestDocument134 pagesVersión Reducida Del Boston Naming Test Tesis Genetica Del TestEduardoCórdovaNo ratings yet
- Tratamientos de Los Trastornos de La Voz MOrrisonDocument311 pagesTratamientos de Los Trastornos de La Voz MOrrisonMarianela Silva100% (5)
- Bactrol e 1110Document2 pagesBactrol e 1110Alejandro Ceron GuerreroNo ratings yet
- Alteraciones Respiratorias Del Recién NacidoDocument1 pageAlteraciones Respiratorias Del Recién NacidoPedroNo ratings yet
- Sesion Educativa de AnemiaDocument5 pagesSesion Educativa de AnemiaAlejandra Navarro PonceNo ratings yet
- GMP Novartis Basel (Valsartan)Document3 pagesGMP Novartis Basel (Valsartan)Aangie EspinosaNo ratings yet
- Cultura, salud y enfermedadDocument28 pagesCultura, salud y enfermedadJavier LabanNo ratings yet
- Factores de Riesgo en Pollos de CarneDocument14 pagesFactores de Riesgo en Pollos de CarneVictoria AguilarNo ratings yet
- Seminario TBCDocument55 pagesSeminario TBCNiko EscuderoNo ratings yet
- Familia HerpesviridaeDocument92 pagesFamilia HerpesviridaerrarNo ratings yet