You might also like
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (121)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (588)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (400)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5794)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1090)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2259)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (345)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- Structural Engineering Formulas Second EditionDocument224 pagesStructural Engineering Formulas Second Editionahmed_60709595194% (33)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (895)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- The Four Types of Gaps and How To Play Them PDFDocument17 pagesThe Four Types of Gaps and How To Play Them PDFshaikiftakar100% (2)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (74)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- Diagnostic Test - Tle8Document2 pagesDiagnostic Test - Tle8rose mae marambaNo ratings yet
- SWAMINATHAN Ajanta RhapsodyDocument227 pagesSWAMINATHAN Ajanta RhapsodyRoberto E. García100% (1)
- Cesars WayDocument20 pagesCesars WayToni TursićNo ratings yet
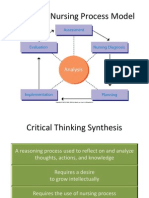
- NUR 104 Nursing Process MY NOTESDocument77 pagesNUR 104 Nursing Process MY NOTESmeanne073100% (1)
- The FlyDocument8 pagesThe FlyDrei Tiam Lacadin100% (1)
- Revised LabDocument18 pagesRevised LabAbu AyemanNo ratings yet
- State of The Art in Research On MicrogridsDocument36 pagesState of The Art in Research On MicrogridsDulal MannaNo ratings yet
- A Software Architecture For The Control of Biomaterials MaintenanceDocument4 pagesA Software Architecture For The Control of Biomaterials MaintenanceCristian ȘtefanNo ratings yet
- Asian RegionalismDocument15 pagesAsian RegionalismLiezel AalaNo ratings yet
- FORM Module IpsDocument10 pagesFORM Module IpsRizalNo ratings yet
- Zoology LAB Scheme of Work 2023 Hsslive HSSDocument7 pagesZoology LAB Scheme of Work 2023 Hsslive HSSspookyvibee666No ratings yet
- Case Study 1 HRM in PandemicDocument2 pagesCase Study 1 HRM in PandemicKristine Dana LabaguisNo ratings yet
- Maharishi Language of Gravity - SoS 27Document3 pagesMaharishi Language of Gravity - SoS 27Prof. MadhavanNo ratings yet
- ASC 2020-21 Questionnaire PDFDocument11 pagesASC 2020-21 Questionnaire PDFShama PhotoNo ratings yet
- Challenges Affecting Devolution PDFDocument94 pagesChallenges Affecting Devolution PDFELIJAH M. OMBEONo ratings yet
- Electrostatics Practice ProblemsDocument4 pagesElectrostatics Practice ProblemsMohammed Aftab AhmedNo ratings yet
- 14 Days of Prayer and FastingDocument40 pages14 Days of Prayer and FastingntsakoramphagoNo ratings yet
- Pamphlet On Arrangement of Springs in Various Casnub Trolleys Fitted On Air Brake Wagon PDFDocument9 pagesPamphlet On Arrangement of Springs in Various Casnub Trolleys Fitted On Air Brake Wagon PDFNiKhil GuPtaNo ratings yet
- Arabian Choice General Trading Co. LLCDocument1 pageArabian Choice General Trading Co. LLCjaanNo ratings yet
- Śāntarak ItaDocument8 pagesŚāntarak ItaÁtilaNo ratings yet
- TRUMPF Marking Laser BrochureDocument48 pagesTRUMPF Marking Laser BrochureKKM SBNo ratings yet
- Veerasaiva Pantha in 12th Century MaharashtraDocument2 pagesVeerasaiva Pantha in 12th Century MaharashtrarathkiraniNo ratings yet
- Estocell - Data Sheet - 14-07-06Document2 pagesEstocell - Data Sheet - 14-07-06LeoRumalaAgusTatarNo ratings yet
- Pediatric Infectious DiseasesDocument3 pagesPediatric Infectious DiseasesAiri FirdausiaNo ratings yet
- BROMINE Safety Handbook - Web FinalDocument110 pagesBROMINE Safety Handbook - Web Finalmonil panchalNo ratings yet
- Math Cad 15Document3 pagesMath Cad 15Kim ChanthanNo ratings yet
- RTRT User GuideDocument324 pagesRTRT User GuideAlae Khaoua100% (3)
- IRAQ Reproductive Maternal, Newborn, Child and Adolescent HealthDocument32 pagesIRAQ Reproductive Maternal, Newborn, Child and Adolescent HealthbejarhasanNo ratings yet