You might also like
- Early Hominin Limb ProportionsDocument20 pagesEarly Hominin Limb Proportions1linNo ratings yet
- De Groote and Humphey 2011 Masa Corp Metatarso1Document8 pagesDe Groote and Humphey 2011 Masa Corp Metatarso1Keila MenaNo ratings yet
- 2012-Richmond & JungersDocument20 pages2012-Richmond & JungersBill JungersNo ratings yet
- Early Homo: Who, When, and WhereDocument21 pagesEarly Homo: Who, When, and WhereAdriana Raluca Ayame SalavastruNo ratings yet
- Journal of Human Evolution: Gabrielle A. Russo, Liza J. ShapiroDocument13 pagesJournal of Human Evolution: Gabrielle A. Russo, Liza J. ShapiroYsNo ratings yet
- Limb Bone Bilateral Asymmetry Variability and Commonality Among Modern HumansDocument16 pagesLimb Bone Bilateral Asymmetry Variability and Commonality Among Modern HumansflorinacretuNo ratings yet
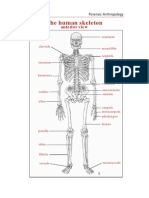
- The Human Skeleton: Anterior ViewDocument0 pagesThe Human Skeleton: Anterior VieweduardoarcticNo ratings yet
- VanderLinden Et Al 2019 Head Turning MorphologiesDocument12 pagesVanderLinden Et Al 2019 Head Turning MorphologiesKo KaNo ratings yet
- Gene Ti Geographic Environmental Correlates Smith Et Al. 2007Document11 pagesGene Ti Geographic Environmental Correlates Smith Et Al. 2007ddrommer1No ratings yet
- Wish Baratz1996Document14 pagesWish Baratz1996smithl25No ratings yet
- Wenner-Gren Foundation For Anthropological Research, The University of Chicago Press Current AnthropologyDocument17 pagesWenner-Gren Foundation For Anthropological Research, The University of Chicago Press Current AnthropologyrandomeoNo ratings yet
- Cranial Variables As Predictors Hominine Body Mass: WordsDocument18 pagesCranial Variables As Predictors Hominine Body Mass: WordsgimenezNo ratings yet
- The Non-Transitions in Human Evolution - On Evolutionists TeDocument6 pagesThe Non-Transitions in Human Evolution - On Evolutionists TepilesarNo ratings yet
- Comparative AnatomyDocument38 pagesComparative AnatomyKimm Charmaine RodriguezNo ratings yet
- Bipedal Locomotion PDFDocument14 pagesBipedal Locomotion PDFCristie SumbillaNo ratings yet
- Pearson 2008 EvAnth Modern Human MorphologyDocument11 pagesPearson 2008 EvAnth Modern Human MorphologyRachid BenNo ratings yet
- Evolution of The Mammalian Neck From Developmental, Morpho-Functional, and Paleontological PerspectivesDocument11 pagesEvolution of The Mammalian Neck From Developmental, Morpho-Functional, and Paleontological PerspectivesKo KaNo ratings yet
- Paleontologia y Sitema ÜticaDocument47 pagesPaleontologia y Sitema Üticaviridana_anahi_2222No ratings yet
- Differentiating Human Bone From Animal Bone A Review of Histological Methods PDFDocument15 pagesDifferentiating Human Bone From Animal Bone A Review of Histological Methods PDFPacifier NothingfaceNo ratings yet
- Hominin Dispersal HistoryDocument34 pagesHominin Dispersal HistoryMarcusNo ratings yet
- Differentiating Human Bone From Animal Bone A Review of Histological MethodsDocument15 pagesDifferentiating Human Bone From Animal Bone A Review of Histological MethodsPacifier NothingfaceNo ratings yet
- Buchholtz Step I en 2009 Ev DevDocument11 pagesBuchholtz Step I en 2009 Ev DevSUKMAWATINo ratings yet
- TIBURONº1Document21 pagesTIBURONº1Jhasmani AmcNo ratings yet
- The University of Chicago Press Wenner-Gren Foundation For Anthropological ResearchDocument22 pagesThe University of Chicago Press Wenner-Gren Foundation For Anthropological ResearchJoel Acuña ZavaletaNo ratings yet
- Brain, Calvarium, Cladistics A New Approach To An Old Question, Who Are Modern Humans and Neandertals - Journal of Human Evolution - 201Document15 pagesBrain, Calvarium, Cladistics A New Approach To An Old Question, Who Are Modern Humans and Neandertals - Journal of Human Evolution - 201Miguel Ángel Badal SalvadorNo ratings yet
- Harmon 2007 - GMM Da Epífise Distal Do FêmurDocument16 pagesHarmon 2007 - GMM Da Epífise Distal Do FêmurTábata ZanescoNo ratings yet
- Limbs in Whales and Limblessness in Other VertebratesDocument14 pagesLimbs in Whales and Limblessness in Other VertebratesJay LeeNo ratings yet
- 2012 Oviraptorosaur Tail Forms and FunctionsDocument47 pages2012 Oviraptorosaur Tail Forms and FunctionsJorge BarrientosNo ratings yet
- Direct evidence of group behavior and locomotion in Homo erectus from 1.5-million-year-old footprintsDocument9 pagesDirect evidence of group behavior and locomotion in Homo erectus from 1.5-million-year-old footprintsvictor muñozNo ratings yet
- Cioclovina Cranium Affinities Early Modern EuropeanDocument15 pagesCioclovina Cranium Affinities Early Modern EuropeanTudor HilaNo ratings yet
- Baab 2016 HerectusneurocranialshapeDocument22 pagesBaab 2016 HerectusneurocranialshapeAwel OumerNo ratings yet
- Healy Et Al. 2015 Ontogenetic Variation in Ramus Form in Neandertals and Modern HumansDocument1 pageHealy Et Al. 2015 Ontogenetic Variation in Ramus Form in Neandertals and Modern Humanschealy21No ratings yet
- Harris Lineas - Nowack 2000Document6 pagesHarris Lineas - Nowack 2000Constanza UrrutiaNo ratings yet
- The Relationship Between Skull Morphology, Masticatory Muscle Force and Cranial Skeletal Deformation During BitingDocument12 pagesThe Relationship Between Skull Morphology, Masticatory Muscle Force and Cranial Skeletal Deformation During BitingMutia KumalasariNo ratings yet
- Ward 2015 AustralopitecinosDocument14 pagesWard 2015 Australopitecinostomascostelafre1410No ratings yet
- Muscle Forces and The Demands of Human Walking: Adam D. Sylvester, Steven G. Lautzenheiser and Patricia Ann KramerDocument10 pagesMuscle Forces and The Demands of Human Walking: Adam D. Sylvester, Steven G. Lautzenheiser and Patricia Ann KramerCarmen Cuerdo del RíoNo ratings yet
- Phil. Trans. R. Soc. B 2010 Lovejoy 3289 99Document12 pagesPhil. Trans. R. Soc. B 2010 Lovejoy 3289 99carlosart26No ratings yet
- Key to the identification and ecology of Cyclopoida (Crustacea, Copepoda) of North America (north of Mexico)From EverandKey to the identification and ecology of Cyclopoida (Crustacea, Copepoda) of North America (north of Mexico)No ratings yet
- Flores Hominid: New Species or Microcephalic Dwarf ?Document23 pagesFlores Hominid: New Species or Microcephalic Dwarf ?ScientificBoyNo ratings yet
- Trinkaus - Neandertal Knees and Ankles A Comment On Miller and GrossDocument2 pagesTrinkaus - Neandertal Knees and Ankles A Comment On Miller and GrossmarziehmarziehNo ratings yet
- Cria de CamelidosDocument22 pagesCria de CamelidosMiranda JosefaNo ratings yet
- The Evolution and Development of Cranial Form in Homo SapiensDocument6 pagesThe Evolution and Development of Cranial Form in Homo SapiensJavier Farias VeraNo ratings yet
- Five Years of H Flores Iens IsDocument13 pagesFive Years of H Flores Iens IsnomadNo ratings yet
- The Anatomical Record - 2009 - Holliday - New Insights Into Dinosaur Jaw Muscle AnatomyDocument20 pagesThe Anatomical Record - 2009 - Holliday - New Insights Into Dinosaur Jaw Muscle AnatomyKidus TarekegnNo ratings yet
- Body elongation in mustelids associated with small size and reduced limbsDocument15 pagesBody elongation in mustelids associated with small size and reduced limbsALISSON JERALDINNE RODRIGUEZ GIRALDONo ratings yet
- Annotated Bibliography For AnthDocument5 pagesAnnotated Bibliography For Anthbaba toundeNo ratings yet
- Joa 228 877Document10 pagesJoa 228 877Kerlida SantosNo ratings yet
- The Evolution of The Upright Posture and Gait-A ReviewDocument23 pagesThe Evolution of The Upright Posture and Gait-A ReviewRicardo Filipe Afonso MangeronaNo ratings yet
- Should Homo Habilis Be Reassigned To Australopithecus?Document3 pagesShould Homo Habilis Be Reassigned To Australopithecus?Pete FlintNo ratings yet
- Hominin Life HistoryDocument32 pagesHominin Life HistorylauraethosNo ratings yet
- Farl Hurlbetal 05 Gator JVP25Document17 pagesFarl Hurlbetal 05 Gator JVP25patelsmita060No ratings yet
- Collar Et Al. 2008Document3 pagesCollar Et Al. 2008Jade CaputoNo ratings yet
- The Study of Skeletal Part Profiles:: An Ambiguous Taphonomic Tool For ZooarchaeologyDocument10 pagesThe Study of Skeletal Part Profiles:: An Ambiguous Taphonomic Tool For ZooarchaeologyProciuc MarianaNo ratings yet
- Wright 2003Document19 pagesWright 2003dario susanoNo ratings yet
- Bailey-2002-The Anatomical Record PDFDocument9 pagesBailey-2002-The Anatomical Record PDFAndres BarriosNo ratings yet
- 2011 On The Evolution of Human Jaws and Teeth A ReviewDocument11 pages2011 On The Evolution of Human Jaws and Teeth A ReviewbaridinoNo ratings yet
- Jen Tree 2007 2Document2 pagesJen Tree 2007 2inditeamNo ratings yet
- Ubelaker. Taphonomic Applications in Forensic AnthropologyDocument14 pagesUbelaker. Taphonomic Applications in Forensic AnthropologyAle Arteaga100% (1)
- Close Encounters of The Prehistoric Kind: OnlineDocument5 pagesClose Encounters of The Prehistoric Kind: Onlinejohnny cartinNo ratings yet
- A Cranium For The Earliest Europeans: Phylogenetic Position of The Hominid From Ceprano, ItalyDocument6 pagesA Cranium For The Earliest Europeans: Phylogenetic Position of The Hominid From Ceprano, Italyjohnny cartinNo ratings yet
- 723 (2010) Hernán A. Burbano,: Science Et AlDocument4 pages723 (2010) Hernán A. Burbano,: Science Et Aljohnny cartinNo ratings yet
- A Draft Sequence of The Neandertal Genome (Sapiens/Neanderthal Interbreeding)Document14 pagesA Draft Sequence of The Neandertal Genome (Sapiens/Neanderthal Interbreeding)José Pedro GomesNo ratings yet
- The First Female Homo Erectus Pelvis, From Gona, Afar, EthiopiaDocument5 pagesThe First Female Homo Erectus Pelvis, From Gona, Afar, Ethiopiajohnny cartinNo ratings yet
- Neanader 2 PDFDocument5 pagesNeanader 2 PDFjohnny cartinNo ratings yet
- Gorillas in CrisisDocument2 pagesGorillas in Crisisjohnny cartinNo ratings yet
- A Cranium For The Earliest Europeans: Phylogenetic Position of The Hominid From Ceprano, ItalyDocument6 pagesA Cranium For The Earliest Europeans: Phylogenetic Position of The Hominid From Ceprano, Italyjohnny cartinNo ratings yet
- Ts17 Gorilla EDocument84 pagesTs17 Gorilla Ejohnny cartinNo ratings yet
- Cell, Vol. 90, 19-30, July 11, 1997, Copyright ©1997 byDocument12 pagesCell, Vol. 90, 19-30, July 11, 1997, Copyright ©1997 byjohnny cartinNo ratings yet
- Proc. Natl. Acad. Sci. USA Vol. 93, Pp. 10852-10854, OctoberDocument3 pagesProc. Natl. Acad. Sci. USA Vol. 93, Pp. 10852-10854, Octoberjohnny cartinNo ratings yet
- Homo Erectus2Document2 pagesHomo Erectus2johnny cartinNo ratings yet
- Inconsistencies in Neanderthal Genomic DNA SequencesDocument5 pagesInconsistencies in Neanderthal Genomic DNA Sequencesjohnny cartinNo ratings yet
- Neanader 2 PDFDocument5 pagesNeanader 2 PDFjohnny cartinNo ratings yet
- Earliest Evidence of Modern Human Life History in North AfricanDocument6 pagesEarliest Evidence of Modern Human Life History in North Africanjohnny cartinNo ratings yet
- Ieidelbergensis 1Document10 pagesIeidelbergensis 1johnny cartinNo ratings yet
- Erg Aster 4Document8 pagesErg Aster 4johnny cartinNo ratings yet
- Homo Erectus1Document4 pagesHomo Erectus1johnny cartinNo ratings yet
- Annali Dell'Università Degli Studi Di Ferrara Museologia Scientifica e NaturalisticaDocument5 pagesAnnali Dell'Università Degli Studi Di Ferrara Museologia Scientifica e Naturalisticajohnny cartinNo ratings yet
- Erg Aster 1Document12 pagesErg Aster 1johnny cartinNo ratings yet
- Homo Erectus4Document18 pagesHomo Erectus4johnny cartinNo ratings yet
- A New Skull of Early Homo From Dmanisi, Georgia AbesalomDocument6 pagesA New Skull of Early Homo From Dmanisi, Georgia Abesalomjohnny cartinNo ratings yet
- Foraging of Homo Ergaster and Australopithecus Boisei in East African EnvironmentsDocument6 pagesForaging of Homo Ergaster and Australopithecus Boisei in East African Environmentsjohnny cartinNo ratings yet
- Controversial Human Ancestor Gets Major Facelift - LiveScienceDocument3 pagesControversial Human Ancestor Gets Major Facelift - LiveSciencejohnny cartinNo ratings yet
- Ann. Zoot. Fennici 28:37/-380 Helsi Nki 19 February 1992Document10 pagesAnn. Zoot. Fennici 28:37/-380 Helsi Nki 19 February 1992johnny cartinNo ratings yet
- American Journal of Physical Anthropology 138:90-100 (2009)Document11 pagesAmerican Journal of Physical Anthropology 138:90-100 (2009)johnny cartinNo ratings yet
- Chapter B The Great Ice AgeDocument2 pagesChapter B The Great Ice Agejohnny cartinNo ratings yet
- Ouranopithecus MacedoniensisDocument27 pagesOuranopithecus Macedoniensisjohnny cartinNo ratings yet
- Journal of Human Evolution 46 (2004) 433-465Document33 pagesJournal of Human Evolution 46 (2004) 433-465johnny cartinNo ratings yet