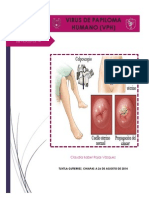
You might also like
- Vacuna contra el Virus del Papiloma Humano: La única vacuna cuestionada incluso por gran parte de la comunidad médicaFrom EverandVacuna contra el Virus del Papiloma Humano: La única vacuna cuestionada incluso por gran parte de la comunidad médicaNo ratings yet
- Monografia VPH.Document5 pagesMonografia VPH.Kevin SánchezNo ratings yet
- Diagnostico Tratamiento y Prevalencia enDocument28 pagesDiagnostico Tratamiento y Prevalencia enkarla.dmcmmNo ratings yet
- Incan Olc 063 3RDocument9 pagesIncan Olc 063 3Rvega_utukku1877No ratings yet
- Virus Del Papiloma HumanoDocument8 pagesVirus Del Papiloma HumanoClaudia RuizNo ratings yet
- Investigacion SSRO MizqueDocument30 pagesInvestigacion SSRO MizqueDi Claros Grágeda0% (1)
- Marco Teorico PreDocument16 pagesMarco Teorico PreMiguel VGNo ratings yet
- Virus Del Papiloma HumanoDocument5 pagesVirus Del Papiloma HumanoSofia RamosNo ratings yet
- Lesiones Premalignas y Malignas de CérvixDocument11 pagesLesiones Premalignas y Malignas de Cérvixapi-19492580No ratings yet
- Resumen de Papiloma HumanoDocument4 pagesResumen de Papiloma HumanoJessica Salmai Cisneros JuárezNo ratings yet
- 5.-Patologia de Cuello Uterino PDFDocument4 pages5.-Patologia de Cuello Uterino PDFGerardo LermaNo ratings yet
- Cáncer CervicouterinoDocument12 pagesCáncer CervicouterinoMarian MirandaNo ratings yet
- Historia Natural de La Enfermedad - EjercicioDocument5 pagesHistoria Natural de La Enfermedad - EjercicioJuan MenaNo ratings yet
- Facultad de Ciencias de La Salud Carrera Profesional de MedicinaDocument16 pagesFacultad de Ciencias de La Salud Carrera Profesional de MedicinaBryam yuri jose Borda sauñeNo ratings yet
- Carcinogenesis Por Virus-Seminario 5Document39 pagesCarcinogenesis Por Virus-Seminario 5Ariana QuenayaNo ratings yet
- Tesis FinalDocument59 pagesTesis FinalJahaziel Gasperin BulbarelaNo ratings yet
- Resumen VPHDocument2 pagesResumen VPHJohnny ArzetaNo ratings yet
- 3RA LECTURA. Sistema Inmunitario y VPHDocument2 pages3RA LECTURA. Sistema Inmunitario y VPHJulisa Sulca RiveraNo ratings yet
- Tipos de InfeccionesDocument11 pagesTipos de InfeccionesANTONIO ANGEL MONTES LEALNo ratings yet
- Protocolo VPHDocument26 pagesProtocolo VPHClaudiaRojasNo ratings yet
- Detección de Linfocitos de Memoria Específicos Contra El Vph-16 en Hombres Y Mujeres Con Vida Sexual ActivaDocument14 pagesDetección de Linfocitos de Memoria Específicos Contra El Vph-16 en Hombres Y Mujeres Con Vida Sexual ActivaJavier YescasNo ratings yet
- Cancer Cervicouterino: Epidemiologia, Historia Natural y Rol Del Virus Papiloma Humano. Perspectivas en Prevencion y TratamientoDocument6 pagesCancer Cervicouterino: Epidemiologia, Historia Natural y Rol Del Virus Papiloma Humano. Perspectivas en Prevencion y TratamientoAlbertoNo ratings yet
- CA de CérvixDocument21 pagesCA de CérvixYanelinEstefanyHernandezCanoNo ratings yet
- Unidad 8 Papilomavirus - VihDocument18 pagesUnidad 8 Papilomavirus - VihLISBETH PANIAGUA GUTIERREZNo ratings yet
- Objetivos Pato 3er ParcialDocument71 pagesObjetivos Pato 3er ParcialElizabeth Evans75No ratings yet
- Virus Piloma HumanoDocument6 pagesVirus Piloma HumanoAlexéiNo ratings yet
- Cáncer de Cuello Uterino-Marco TeoricoDocument2 pagesCáncer de Cuello Uterino-Marco TeoricoWillMonasHuertas100% (4)
- PapilomasDocument8 pagesPapilomasClaudia Cochachin ChuicaNo ratings yet
- 11virus Papoloma Humano, Neoplasia Cervical Cancer de Vulva Cancer de VaginaDocument35 pages11virus Papoloma Humano, Neoplasia Cervical Cancer de Vulva Cancer de VaginaEdwing Sedano GamarraNo ratings yet
- A.R.-infección Por El Virus Del Papiloma HumanoDocument13 pagesA.R.-infección Por El Virus Del Papiloma HumanoMelanie Muñoz RuizNo ratings yet
- 431-Texto Del Artículo-769-2-10-20210407Document17 pages431-Texto Del Artículo-769-2-10-20210407Brenda Hernández TrejoNo ratings yet
- Articulo PPT Vih SidaDocument12 pagesArticulo PPT Vih SidaErick Liñan AlarconNo ratings yet
- Ensayo VPHDocument4 pagesEnsayo VPHDaniela DurónNo ratings yet
- Que Ès EtiologìaDocument7 pagesQue Ès EtiologìaJenifer Fresia Zegarra RodriguezNo ratings yet
- Papiloma Humano Trabajo OficialDocument8 pagesPapiloma Humano Trabajo OficialVicenteRamosMamaniNo ratings yet
- DiferenciaciónDocument7 pagesDiferenciaciónkrisel andreina velasquez meignenNo ratings yet
- CITOMEGALOVIRUSDocument7 pagesCITOMEGALOVIRUSJosé Luis Luviano EscobarNo ratings yet
- RMD 191 FDocument20 pagesRMD 191 FJoel RodríguezNo ratings yet
- Virus Del Papiloma HumanoDocument6 pagesVirus Del Papiloma HumanoJosé Fernando Salgado MorenoNo ratings yet
- PapovavirusDocument3 pagesPapovavirusAnonymous TN4XtXvNo ratings yet
- Ebook Historia Natural Del CancerDocument6 pagesEbook Historia Natural Del CancerDai García de CitorushtcNo ratings yet
- 28 Cancer de Cuello UterinoDocument14 pages28 Cancer de Cuello UterinoSAULNo ratings yet
- Ficha de Estudio - Virologia - VPHDocument3 pagesFicha de Estudio - Virologia - VPHkevinNo ratings yet
- Informe II PatologiaDocument117 pagesInforme II PatologiaDanyelo Sandoval CerdánNo ratings yet
- Investigación VPH - MicrobiologiaDocument17 pagesInvestigación VPH - MicrobiologiaRonald Celi CastilloNo ratings yet
- Cancer de Cervix1Document8 pagesCancer de Cervix1José Miguel Santana HerreraNo ratings yet
- Cancer Cervico UterinoDocument15 pagesCancer Cervico UterinoErick Ariel Camacho AguilarNo ratings yet
- IntroduccionDocument14 pagesIntroduccionGABRIELA ESTEFANIA REYES PINEDANo ratings yet
- Infección Por Virus Del Papiloma Humano (VPH) - Vacunas Profilácticas. Indicaciones.Document20 pagesInfección Por Virus Del Papiloma Humano (VPH) - Vacunas Profilácticas. Indicaciones.Conrad SalzNo ratings yet
- VPH SemDocument15 pagesVPH SemMayra Alejandra GonzalesNo ratings yet
- P0liomavirus y PapilomavirusDocument35 pagesP0liomavirus y PapilomavirusNelidaMamaniNo ratings yet
- Susceptibilidad Genética Frente Al Cáncer Bucal Por Infección Del Virus Del Papiloma HumanoDocument7 pagesSusceptibilidad Genética Frente Al Cáncer Bucal Por Infección Del Virus Del Papiloma HumanoSELENE SHARMILA SUMOSO ORDONEZNo ratings yet
- 27 VihDocument5 pages27 VihJosé Pablo Perea RoveloNo ratings yet
- ARTÍCULO - Mala Protección en Relaciones Sexuales Como Detonante de Cancer CervicalDocument2 pagesARTÍCULO - Mala Protección en Relaciones Sexuales Como Detonante de Cancer CervicalDignorah ParedesNo ratings yet
- Carina Alarcon Vila 2Document31 pagesCarina Alarcon Vila 2samNo ratings yet
- Unidad 8 Papilomavirus - VihDocument22 pagesUnidad 8 Papilomavirus - VihPedro Nascimento da SilvaNo ratings yet
- Cáncer Cérvico Uterino: Prevención y Tratamiento Cervical Uterine Cancer: Prevention and TreatmentDocument9 pagesCáncer Cérvico Uterino: Prevención y Tratamiento Cervical Uterine Cancer: Prevention and TreatmentBelén SifuentesNo ratings yet
- Virus Del Papiloma Humano VPH Cito3erparcialDocument5 pagesVirus Del Papiloma Humano VPH Cito3erparcialRosario Calani CanaviriNo ratings yet
- PREVENCION PRIMARIA CANCER CERVICO UTERINO ContentServer PDFDocument11 pagesPREVENCION PRIMARIA CANCER CERVICO UTERINO ContentServer PDFJorge CmpNo ratings yet
- Tabla Conversion CompletaDocument12 pagesTabla Conversion CompletaPeter Harold El Tigre86% (64)
- Conflicto en La ComunicacionDocument7 pagesConflicto en La Comunicacionhector_212No ratings yet
- Libro Bitacora de La UtopiaDocument100 pagesLibro Bitacora de La UtopiaCamilo cantorNo ratings yet
- Clasificacion NumerosDocument1 pageClasificacion NumerosEdwin MhdezNo ratings yet
- Fisiologia Del EmbarazoDocument23 pagesFisiologia Del EmbarazoEdwin MhdezNo ratings yet
- GeneracionDocument3 pagesGeneracionEdwin MhdezNo ratings yet
- Plaga PrimariaDocument10 pagesPlaga PrimariaarrebatamientoNo ratings yet
- Manual de Operacion y Mantenimiento PtarDocument40 pagesManual de Operacion y Mantenimiento PtarEdna CepedaNo ratings yet
- Plantilla JouleDocument8 pagesPlantilla JouleAurelio CanazaNo ratings yet
- Dictamen de Lesiones PersonalesDocument73 pagesDictamen de Lesiones PersonalesSofia MontalvoNo ratings yet
- 04 - Prueba de Avance 04 - Biología - Química - CV - RespuestasDocument5 pages04 - Prueba de Avance 04 - Biología - Química - CV - RespuestasDiego Andres Garcia MurciaNo ratings yet
- 5 Catalogo de Patologias CIE 10Document136 pages5 Catalogo de Patologias CIE 10SSTNo ratings yet
- Virus Papiloma HumanoDocument6 pagesVirus Papiloma HumanoAnyelina BlancoNo ratings yet
- Biologia SuperiorDocument98 pagesBiologia SuperiorMARIA MITESNo ratings yet
- Diagnóstico Microbiológico de Las Infecciones Del Tracto Respiratorio SuperiorDocument29 pagesDiagnóstico Microbiológico de Las Infecciones Del Tracto Respiratorio SuperiorMagali YuruhanNo ratings yet
- 3.reinos de Seres VivosDocument21 pages3.reinos de Seres VivosMAR�A DOLORES HERRERA POYATOSNo ratings yet
- Cuadernillo N°4 (Ciencias 7°) ReproduccionDocument31 pagesCuadernillo N°4 (Ciencias 7°) ReproduccionFrancisco Osvaldo Hernández LópezNo ratings yet
- Ciencias Naturales 9no U 1-5Document40 pagesCiencias Naturales 9no U 1-5Alex RodríguezNo ratings yet
- Catalogo SanitarioDocument4 pagesCatalogo Sanitarioelio carboNo ratings yet
- El Virus de La Inmunodeficiencia HumanaDocument31 pagesEl Virus de La Inmunodeficiencia HumanaRosanna LebronNo ratings yet
- Huésped U HospederoDocument18 pagesHuésped U HospederocarlosNo ratings yet
- Examen Primer Parcial InfecciosasDocument11 pagesExamen Primer Parcial InfecciosasPaula GarroteNo ratings yet
- Tarea de Genetica Carlos Palacios Lindo 85111Document3 pagesTarea de Genetica Carlos Palacios Lindo 85111Carlos Enrique PalaciosNo ratings yet
- Control Biologico Una Forma Sustentable de Control de PlagasDocument8 pagesControl Biologico Una Forma Sustentable de Control de PlagasJulian LeonardoNo ratings yet
- "Arzobispo Loayza": Guia de Procedimiento para Enfermeria TecnicaDocument22 pages"Arzobispo Loayza": Guia de Procedimiento para Enfermeria TecnicaBladimir F J MendezNo ratings yet
- Clase 01. Morfologia MicrobianaDocument48 pagesClase 01. Morfologia MicrobianaCristian Pincheira ArayaNo ratings yet
- Enfermedades FebrilesDocument48 pagesEnfermedades FebrilesGeraldineAlcantaraMalaverNo ratings yet
- Paper Ajustado 2023Document24 pagesPaper Ajustado 2023Natalia Acosta OliveraNo ratings yet
- RevistadeOccidente Septiembre2018 M - Isabel-Porras-GalloDocument12 pagesRevistadeOccidente Septiembre2018 M - Isabel-Porras-GalloleanfilmsNo ratings yet
- Escenarios para El Futuro de La Tecnología y El Desarrollo Internacional en PDFDocument32 pagesEscenarios para El Futuro de La Tecnología y El Desarrollo Internacional en PDFekkexxe100% (1)
- Epstein Barr PDFDocument16 pagesEpstein Barr PDFCuquis Vargas BravoNo ratings yet
- Guia de Biologia Ceneval 286Document25 pagesGuia de Biologia Ceneval 286Anonymous Y6r5JnQNo ratings yet
- Act-Desarr 5Document134 pagesAct-Desarr 5Max MarNo ratings yet
- Manual de Laboratorio VirologiaDocument67 pagesManual de Laboratorio VirologiaJohn ChavezNo ratings yet
- ESTUDIO DE LA PANDEMIA y ANÁLISIS CIENTÍFICO INDEPENDIENTE Dr. Sergio J. Pérez OliveroDocument2,290 pagesESTUDIO DE LA PANDEMIA y ANÁLISIS CIENTÍFICO INDEPENDIENTE Dr. Sergio J. Pérez OliveroYankaNo ratings yet
- Encefalitis BC TES TMP 2482Document24 pagesEncefalitis BC TES TMP 2482OrganicAgriculturePharmacologyDosologyHersil PfizerNo ratings yet