You might also like
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (895)
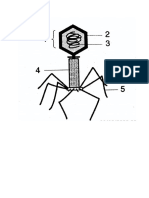
- Virus GMBRDocument4 pagesVirus GMBRTutik WulandariNo ratings yet
- PENGARUH PEMBERIAN Curcuma Domestica Terhadap Gambaran Histologi Hepar Mencit Balb/C Yang Diberi ParasetamolDocument30 pagesPENGARUH PEMBERIAN Curcuma Domestica Terhadap Gambaran Histologi Hepar Mencit Balb/C Yang Diberi ParasetamolTutik WulandariNo ratings yet
- 11 0903 Anataomy LILIUM Am0505Document6 pages11 0903 Anataomy LILIUM Am0505Tutik WulandariNo ratings yet
- Bauhinia PurpureaDocument3 pagesBauhinia PurpureaTutik WulandariNo ratings yet
- Antibiotics: Biotechnology IIDocument11 pagesAntibiotics: Biotechnology IITutik WulandariNo ratings yet
- Biji Keras 1-2 PDFDocument11 pagesBiji Keras 1-2 PDFTutik WulandariNo ratings yet
- 7 13v3i4 3Document6 pages7 13v3i4 3Tutik WulandariNo ratings yet
- Materi GenetikaDocument30 pagesMateri GenetikaRizka Eka Apc100% (1)
- Classis AvesDocument54 pagesClassis AvesTutik WulandariNo ratings yet
- Classis AvesDocument54 pagesClassis AvesTutik WulandariNo ratings yet
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5794)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (399)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (588)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2259)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (73)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (344)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (121)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- Science Grade 10 (Exam Prep)Document6 pagesScience Grade 10 (Exam Prep)Venice Solver100% (3)
- List of Bird Sanctuaries in India (State-Wise)Document6 pagesList of Bird Sanctuaries in India (State-Wise)VISHRUTH.S. GOWDANo ratings yet
- A Tall Order - Cooling Dubai's Burj Khalifa: FeatureDocument2 pagesA Tall Order - Cooling Dubai's Burj Khalifa: FeatureMohsin KhanNo ratings yet
- General Characteristics of Phonemes: Aspects of Speech SoundsDocument8 pagesGeneral Characteristics of Phonemes: Aspects of Speech SoundsElina EkimovaNo ratings yet
- Ebook Stackoverflow For ItextDocument336 pagesEbook Stackoverflow For ItextAnonymous cZTeTlkag9No ratings yet
- Thompson, Damon - Create A Servitor - How To Create A Servitor and Use The Power of Thought FormsDocument49 pagesThompson, Damon - Create A Servitor - How To Create A Servitor and Use The Power of Thought FormsMike Cedersköld100% (5)
- Department of Education: Republic of The PhilippinesDocument1 pageDepartment of Education: Republic of The PhilippinesKlaribelle VillaceranNo ratings yet
- ITCNASIA23 - Visitor GuideDocument24 pagesITCNASIA23 - Visitor Guideibrahim shabbirNo ratings yet
- Operator'S Manual PM20X-X-X-BXX: 2" Diaphragm PumpDocument12 pagesOperator'S Manual PM20X-X-X-BXX: 2" Diaphragm PumpOmar TadeoNo ratings yet
- R, Axn: Housingand RegulatoryDocument5 pagesR, Axn: Housingand RegulatoryAce RamosoNo ratings yet
- Multi-Media Approach To Teaching-LearningDocument8 pagesMulti-Media Approach To Teaching-LearningswethashakiNo ratings yet
- Iroquois Clothes and WampumDocument3 pagesIroquois Clothes and Wampumapi-254323856No ratings yet
- SAP HCM Case StudyDocument17 pagesSAP HCM Case StudyRafidaFatimatuzzahraNo ratings yet
- IGCSE 0408 Unseen Poem QuestionsDocument5 pagesIGCSE 0408 Unseen Poem QuestionsMenon HariNo ratings yet
- Distance SortDocument6 pagesDistance SortAI Coordinator - CSC JournalsNo ratings yet
- L5T-112 Manual - 2007 - Issue 1.1 PDFDocument16 pagesL5T-112 Manual - 2007 - Issue 1.1 PDFfluidaimaginacionNo ratings yet
- GladioDocument28 pagesGladioPedro Navarro SeguraNo ratings yet
- دور أخلاقيات الأعمال في تحسين أداء المنظماتDocument14 pagesدور أخلاقيات الأعمال في تحسين أداء المنظماتChaima LaifaNo ratings yet
- Vtoris 100% Clean Paypal Transfer Guide 2015Document8 pagesVtoris 100% Clean Paypal Transfer Guide 2015Sean FrohmanNo ratings yet
- Under Suitable Conditions, Butane, C: © OCR 2022. You May Photocopy ThisDocument13 pagesUnder Suitable Conditions, Butane, C: © OCR 2022. You May Photocopy ThisMahmud RahmanNo ratings yet
- Ericsson For Sale From Powerstorm 4SA03071242Document8 pagesEricsson For Sale From Powerstorm 4SA03071242wd3esaNo ratings yet
- Medical Equipment Quality Assurance For Healthcare FacilitiesDocument5 pagesMedical Equipment Quality Assurance For Healthcare FacilitiesJorge LopezNo ratings yet
- Chandigarh Distilers N BotlersDocument3 pagesChandigarh Distilers N BotlersNipun GargNo ratings yet
- NZ2016SH (32k) - e - NSC5026D 3.3V +100ppmDocument2 pagesNZ2016SH (32k) - e - NSC5026D 3.3V +100ppmDumarronNo ratings yet
- RseDocument60 pagesRseH S Vishwanath ShastryNo ratings yet
- Adding and Subtracting FractionsDocument4 pagesAdding and Subtracting Fractionsapi-508898016No ratings yet
- PRINCIPLES OF TEACHING NotesDocument24 pagesPRINCIPLES OF TEACHING NotesHOLLY MARIE PALANGAN100% (2)
- 4D Beijing (Muslim) CHINA MATTA Fair PackageDocument1 page4D Beijing (Muslim) CHINA MATTA Fair PackageSedunia TravelNo ratings yet
- Poka-Yoke or Mistake Proofing: Historical Evolution.Document5 pagesPoka-Yoke or Mistake Proofing: Historical Evolution.Harris ChackoNo ratings yet
- Windows Insider ProgramDocument10 pagesWindows Insider ProgramVasileBurcuNo ratings yet