You might also like
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5794)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1090)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (894)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (587)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (344)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (119)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (399)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2219)
- A Trip To The Beach Case StudyDocument3 pagesA Trip To The Beach Case StudynadhraNo ratings yet
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (265)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- Self-I-Dentity Through Ho'oponoponoDocument5 pagesSelf-I-Dentity Through Ho'oponoponoTanya McCracken100% (1)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (73)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- Princeton Guide To EcologyDocument3 pagesPrinceton Guide To EcologyAngie ReyesNo ratings yet
- UtruDocument67 pagesUtruTezadeLicentaNo ratings yet
- Investigations CriminalDocument88 pagesInvestigations CriminalTezadeLicentaNo ratings yet
- Momi Documentation: Release 2.0.1Document29 pagesMomi Documentation: Release 2.0.1TezadeLicentaNo ratings yet
- WEF Human Capital Report 2015 PDFDocument319 pagesWEF Human Capital Report 2015 PDFTezadeLicentaNo ratings yet
- PoliticaDocument14 pagesPoliticaTezadeLicentaNo ratings yet
- Uha Bcde GHJ: Air Assisted Lances Single Orifice NozzlesDocument2 pagesUha Bcde GHJ: Air Assisted Lances Single Orifice NozzlesraamonikNo ratings yet
- ManaDocument14 pagesManaTezadeLicentaNo ratings yet
- SnailDocument25 pagesSnailBolarinwaNo ratings yet
- PopaDocument292 pagesPopaTezadeLicentaNo ratings yet
- Koker ThermostatDocument18 pagesKoker ThermostatTezadeLicentaNo ratings yet
- LolarDocument6 pagesLolarTezadeLicentaNo ratings yet
- Bittersweet Cookies. Some Security and Privacy ConsiderationsDocument16 pagesBittersweet Cookies. Some Security and Privacy ConsiderationsStefano BendandiNo ratings yet
- High-speed electronic eyelet buttonholer with 2,500 spmDocument6 pagesHigh-speed electronic eyelet buttonholer with 2,500 spmTezadeLicentaNo ratings yet
- HororDocument11 pagesHororTezadeLicentaNo ratings yet
- DM StatDocument7 pagesDM StatlxangelusxlNo ratings yet
- GugDocument56 pagesGugTezadeLicentaNo ratings yet
- FuturesDocument16 pagesFuturesTezadeLicentaNo ratings yet
- LolaDocument12 pagesLolaTezadeLicentaNo ratings yet
- ! Sustainable Energy Synopsis10Document10 pages! Sustainable Energy Synopsis10quikjipNo ratings yet
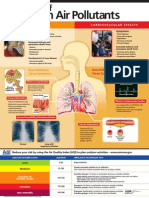
- Effects Of: Common Air PollutantsDocument1 pageEffects Of: Common Air PollutantsTezadeLicentaNo ratings yet
- KiloDocument2 pagesKiloTezadeLicentaNo ratings yet
- Bernhard Jucker: Head of Power Products Division Member of The Group Executive Committee of ABB LTD, SwitzerlandDocument1 pageBernhard Jucker: Head of Power Products Division Member of The Group Executive Committee of ABB LTD, SwitzerlandTezadeLicentaNo ratings yet
- Eadweard MuybridgeDocument9 pagesEadweard MuybridgeTezadeLicentaNo ratings yet
- SchemaDocument6 pagesSchemaTezadeLicentaNo ratings yet
- How Science WorksDocument39 pagesHow Science Worksdhani_isNo ratings yet
- FasDocument6 pagesFasTezadeLicentaNo ratings yet
- Karlos CastanDocument3 pagesKarlos CastanTezadeLicentaNo ratings yet
- Textbook of Wild and Zoo AnimalsDocument247 pagesTextbook of Wild and Zoo Animalsprolixe108100% (3)
- EcosystemDocument35 pagesEcosystemBibek RoyNo ratings yet
- Earth & Life Science: Quarter 1 - Module 7Document19 pagesEarth & Life Science: Quarter 1 - Module 7Jofan Kyle TvmcNo ratings yet
- Inigenous Knowledge and Environmental Conservation in East AfricaDocument47 pagesInigenous Knowledge and Environmental Conservation in East AfricaAdonias GeraldNo ratings yet
- Food Chain and Food WebDocument12 pagesFood Chain and Food Websriyoga100% (2)
- Ben Block Presentation To USAIDDocument23 pagesBen Block Presentation To USAIDBueyBenNo ratings yet
- Biotic Factors DefinitionDocument12 pagesBiotic Factors DefinitionNairdax Navie Ziur OzilacNo ratings yet
- Afghan Breckle FloraDocument8 pagesAfghan Breckle FloraRandy PanggabeanNo ratings yet
- NEET UG Biology Organism and PopulationDocument15 pagesNEET UG Biology Organism and PopulationmotivationTechNo ratings yet
- Project Tiger Wildlife Conservation Saves Endangered SpeciesDocument6 pagesProject Tiger Wildlife Conservation Saves Endangered SpeciesNirja palNo ratings yet
- NON-CONVENTIONAL ENERGY RESOURCES TEXTBOOKDocument6 pagesNON-CONVENTIONAL ENERGY RESOURCES TEXTBOOKFatima MirNo ratings yet
- 001 - Capitulo 8 Whitmore (1) para TraducirDocument30 pages001 - Capitulo 8 Whitmore (1) para TraducirMaria joseNo ratings yet
- Women's Role in Environmental Protection in KarnatakaDocument2 pagesWomen's Role in Environmental Protection in KarnatakaNithyananda PatelNo ratings yet
- Kelimpahan Dan Pola Pertumbuhan Kepiting Bakau ScyDocument8 pagesKelimpahan Dan Pola Pertumbuhan Kepiting Bakau ScyNelsi Antonius KalaNo ratings yet
- Amazon River Fascinating FactsDocument13 pagesAmazon River Fascinating FactsvsnpradeepNo ratings yet
- Brg1 AIO More Prac 4Document4 pagesBrg1 AIO More Prac 4Abraham Ortega MorenoNo ratings yet
- Plants and Animals Living TogetherDocument14 pagesPlants and Animals Living TogetherMohamed Mahmoud Mobarak100% (1)
- Leafscale Gulper Shark ST FactsheetDocument5 pagesLeafscale Gulper Shark ST FactsheetLa Ode Khairum MastuNo ratings yet
- Ecological Restoration Source BookDocument109 pagesEcological Restoration Source Bookpalani100% (2)
- Evelyn Thayer Chace Award: Keynote Speaker: John Flicker, President, National Audubon SocietyDocument8 pagesEvelyn Thayer Chace Award: Keynote Speaker: John Flicker, President, National Audubon SocietyAudubonSCNo ratings yet
- Final Chapter 1 - 2 - UnDocument10 pagesFinal Chapter 1 - 2 - UnJoshua PincaNo ratings yet
- Fascinating Facts About MadagascarDocument2 pagesFascinating Facts About MadagascarIrene OriarewoNo ratings yet
- Scottish National Heritage - The Nature of Scotland SpringSummer2015Issue - 21Document35 pagesScottish National Heritage - The Nature of Scotland SpringSummer2015Issue - 21ThomasEngstNo ratings yet
- Assessing Benefits and Costs of Urban ForestsDocument8 pagesAssessing Benefits and Costs of Urban ForestsdmbgddNo ratings yet
- Science Power 10 Chapter Black Line Master PDFDocument39 pagesScience Power 10 Chapter Black Line Master PDFgiri_sundar5930No ratings yet
- Week 1 - Overview of Environmental Management 1Document39 pagesWeek 1 - Overview of Environmental Management 1Joshua NemiNo ratings yet
- ESP3 SEA Synthesis Study April 2013 Final Report ENG Rev0Document126 pagesESP3 SEA Synthesis Study April 2013 Final Report ENG Rev0AhsanaRiskaArifNo ratings yet
- Atlas of Biogeography and Biodiversity of Indiginous Domestic and Wildlife of EthiopiaDocument70 pagesAtlas of Biogeography and Biodiversity of Indiginous Domestic and Wildlife of EthiopiaMelaku Tefera100% (2)