You might also like
- Molecular Genetic Analysis and Regulation of Aflatoxin BiosynthesisDocument11 pagesMolecular Genetic Analysis and Regulation of Aflatoxin BiosynthesisconcatamerNo ratings yet
- Bacterial Gene Aldoketo Reductase Gene CloningDocument8 pagesBacterial Gene Aldoketo Reductase Gene CloningconcatamerNo ratings yet
- Azokeratin ProtocolDocument8 pagesAzokeratin ProtocolconcatamerNo ratings yet
- Acetone Precipitation of Proteins - TECHNICAL RESOURCEDocument2 pagesAcetone Precipitation of Proteins - TECHNICAL RESOURCENathanna MateusNo ratings yet
- Comparison of Protein Precipitation Methods For Sample PreparationDocument4 pagesComparison of Protein Precipitation Methods For Sample PreparationconcatamerNo ratings yet
- Mouse Liver Glutathione S-Transferase Lsoenzyme ActivityDocument10 pagesMouse Liver Glutathione S-Transferase Lsoenzyme ActivityconcatamerNo ratings yet
- Chryseobacterium Library Construction: Restriction Endonuclease RangeDocument2 pagesChryseobacterium Library Construction: Restriction Endonuclease RangeconcatamerNo ratings yet
- For Seminar Molecular Basis of Aflatoxin-Induced MutagenesisDocument8 pagesFor Seminar Molecular Basis of Aflatoxin-Induced MutagenesisconcatamerNo ratings yet
- Transposon LibraryDocument9 pagesTransposon LibraryconcatamerNo ratings yet
- Research OrientationDocument8 pagesResearch OrientationconcatamerNo ratings yet
- Degradation of Aflatoxin B1 by RhodococcusDocument7 pagesDegradation of Aflatoxin B1 by RhodococcusconcatamerNo ratings yet
- Probiotis and Mucus BarrierDocument13 pagesProbiotis and Mucus BarrierconcatamerNo ratings yet
- Xenobiotic Biotransformation Basic ConceptsDocument61 pagesXenobiotic Biotransformation Basic Conceptsconcatamer100% (1)
- Chemistry and Biology of Afb1Document11 pagesChemistry and Biology of Afb1concatamerNo ratings yet
- ASP13 (1) 2014 03-18 MaxDocument16 pagesASP13 (1) 2014 03-18 MaxconcatamerNo ratings yet
- What Is PhilosophyDocument100 pagesWhat Is PhilosophyconcatamerNo ratings yet
- A Rapid Aflatoxin B ELISA: Development and Validation With Reduced Matrix Effects For Peanuts, Corn, Pistachio, and SoybeansDocument10 pagesA Rapid Aflatoxin B ELISA: Development and Validation With Reduced Matrix Effects For Peanuts, Corn, Pistachio, and SoybeansconcatamerNo ratings yet
- Choudhary KeraDocument8 pagesChoudhary KeraconcatamerNo ratings yet
- What Is PhilosophyDocument100 pagesWhat Is PhilosophyconcatamerNo ratings yet
- Degradation of Aflatoxin B1 During The Fermentation of Alcoholic BeveragesDocument11 pagesDegradation of Aflatoxin B1 During The Fermentation of Alcoholic BeveragesconcatamerNo ratings yet
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5794)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (890)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (399)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (344)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (587)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (73)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (265)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1090)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2219)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (119)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- Catalogo Completo Cooper LightingDocument130 pagesCatalogo Completo Cooper LightingWilliam Dante Mescco SaireNo ratings yet
- INTRODUCTION TO COMPASS SURVEYyDocument13 pagesINTRODUCTION TO COMPASS SURVEYyCristiano RonaldoNo ratings yet
- Aggregate PresentationDocument34 pagesAggregate PresentationRahsaan KirtonNo ratings yet
- Mediana Ika Syafirina HDocument6 pagesMediana Ika Syafirina HChristina MargarethaNo ratings yet
- Lab Report 3Document8 pagesLab Report 3sagarchawlaNo ratings yet
- 内容1-HL revision guideDocument231 pages内容1-HL revision guide2193693752No ratings yet
- INSUPOLY 160 PES 4.5 KG Slates Finish-3227120316Document1 pageINSUPOLY 160 PES 4.5 KG Slates Finish-3227120316Jesus Endara condoriNo ratings yet
- Bandstructure Lab FTUG EdtDocument42 pagesBandstructure Lab FTUG EdtAmol HNo ratings yet
- B.Tech. (5 Semester Mechanical) TRIBOLOGY (MEPE-17)Document5 pagesB.Tech. (5 Semester Mechanical) TRIBOLOGY (MEPE-17)varunNo ratings yet
- Numerical Model TurnerDocument696 pagesNumerical Model TurnerChandan Kumar Ray100% (1)
- Use of Surfactants in Metal Cutting FluidsDocument24 pagesUse of Surfactants in Metal Cutting FluidsLen SurbanNo ratings yet
- Environmental Technology & Innovation: Meenal Gupta, Harsh Gupta, D.S. KharatDocument11 pagesEnvironmental Technology & Innovation: Meenal Gupta, Harsh Gupta, D.S. KharatBayu Pramana PutraNo ratings yet
- Proposal FormatDocument21 pagesProposal Formatግሩም ግብርከNo ratings yet
- Elements of Potential Theory: Overview: M U GM/R G M R R G M R N U X U U G M - PDocument15 pagesElements of Potential Theory: Overview: M U GM/R G M R R G M R N U X U U G M - PfucsomoNo ratings yet
- download-Physics-A-level-Notes-Edexcel-IAL-Unit-4-Detailed-4.3. Further MechanicsDocument11 pagesdownload-Physics-A-level-Notes-Edexcel-IAL-Unit-4-Detailed-4.3. Further MechanicsTowsif HassanNo ratings yet
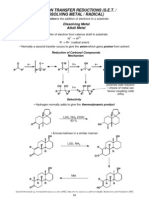
- Metal ReductionDocument11 pagesMetal Reductiondeepthimahanthi sabithaNo ratings yet
- 08 HydroprocessingDocument38 pages08 HydroprocessingrciographyNo ratings yet
- Kumpulan Soal Bioenergetika TugasDocument19 pagesKumpulan Soal Bioenergetika TugasAnika Kunthi HutamiNo ratings yet
- Ideal Fluid Flow Engineering PDFDocument55 pagesIdeal Fluid Flow Engineering PDFsarmadNo ratings yet
- TD RDMDocument3 pagesTD RDMIbtissam LachguerNo ratings yet
- Bearing Spark SiapDocument5 pagesBearing Spark SiapShamsul AimanNo ratings yet
- api 510 اسئلة متنوعة لكل الكورسDocument42 pagesapi 510 اسئلة متنوعة لكل الكورسhussam ghiathNo ratings yet
- Asdadfwassfbdfbdsbfdfsb PDFDocument6 pagesAsdadfwassfbdfbdsbfdfsb PDFpNo ratings yet
- US8178353 - Method For Determination of Polymer Concentration in Water SystemsDocument15 pagesUS8178353 - Method For Determination of Polymer Concentration in Water SystemsKhanh Nguyen TuanNo ratings yet
- ASTM A295-98 (Obsolete)Document2 pagesASTM A295-98 (Obsolete)techietaddyNo ratings yet
- The Heat of Neutralization of Acetic Acid 1Document4 pagesThe Heat of Neutralization of Acetic Acid 1JjjjNo ratings yet
- Bio IGCSE DLP 7Document4 pagesBio IGCSE DLP 7ynr.ermiyasNo ratings yet
- E 155 - 95 - Rte1ns1sruq - PDFDocument6 pagesE 155 - 95 - Rte1ns1sruq - PDFLeón SuárezNo ratings yet
- Chemical Waste Disposal of NEUSTDocument2 pagesChemical Waste Disposal of NEUSTRoniel Urbino GamboaNo ratings yet
- Alloyed: Case-Hardening Steels RM16 - RM16PBDocument3 pagesAlloyed: Case-Hardening Steels RM16 - RM16PBShubhangi ShindeNo ratings yet