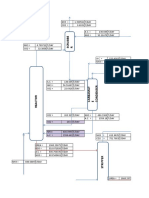
You might also like
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5794)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (895)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (400)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (588)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (74)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (344)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2259)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1090)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (121)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- Octyl Methoxycinnamate: Kyowa Hakko Europe GMBH Kyowa Hakko Europe GMBHDocument4 pagesOctyl Methoxycinnamate: Kyowa Hakko Europe GMBH Kyowa Hakko Europe GMBHDeriven TewengNo ratings yet
- Microbe Deg of PFASDocument25 pagesMicrobe Deg of PFASBJ BIOLOGYNo ratings yet
- Paint School: Training Course HandoutsDocument71 pagesPaint School: Training Course HandoutsShaaban Hamed93% (14)
- Unit 9Document15 pagesUnit 9KOLA AVINASH RAI 218001273No ratings yet
- Why Does Asparagus Make Your Pee Smell PDFDocument145 pagesWhy Does Asparagus Make Your Pee Smell PDFhsrazadNo ratings yet
- United Initiators Mutli Component Peroxides MEKP Is Not MEKP NoroxDocument6 pagesUnited Initiators Mutli Component Peroxides MEKP Is Not MEKP NoroxvasucristalNo ratings yet
- Smart Compost 1Document5 pagesSmart Compost 1YATHISH M GNo ratings yet
- Phys - Mcqs - Endocrine - Gastroenterology 123Document41 pagesPhys - Mcqs - Endocrine - Gastroenterology 123Syeda AlizaNo ratings yet
- Asam Arakhidonat STZDocument6 pagesAsam Arakhidonat STZAlmas TNo ratings yet
- Annex III, Last Update: 20/01/2023: Page 1/91Document91 pagesAnnex III, Last Update: 20/01/2023: Page 1/91Victoria Aguirre MarinNo ratings yet
- Biology EUEE 2013 (14) - 151269132021Document14 pagesBiology EUEE 2013 (14) - 151269132021NebilahNo ratings yet
- Barzaghi General EngDocument12 pagesBarzaghi General Engalberto_71No ratings yet
- A Review of General ChemistryDocument13 pagesA Review of General ChemistryValerie Mae Librero AreñoNo ratings yet
- Chemicals and Fertilizers Industry: Group-3Document11 pagesChemicals and Fertilizers Industry: Group-3Kavya DaraNo ratings yet
- 35 International Chemistry Olympiad: Student Name: Student CodeDocument14 pages35 International Chemistry Olympiad: Student Name: Student CodeLê Hoàng MinhNo ratings yet
- Ulman Part 5Document1,000 pagesUlman Part 5Yana RahmadaniNo ratings yet
- 03 - 05 Transcription and Translation AssignmentDocument2 pages03 - 05 Transcription and Translation AssignmentKeara RaleighNo ratings yet
- New Price List 13 Sept 2022 - Rucika (Pipa, Fitting, Lem) Web - FinalDocument13 pagesNew Price List 13 Sept 2022 - Rucika (Pipa, Fitting, Lem) Web - FinalMaulana IbrahimNo ratings yet
- Headspace Analysis Journal ArticleDocument5 pagesHeadspace Analysis Journal ArticleChristopher GrahamNo ratings yet
- Science11 - Q2 - Mod1 - GeneralBiology1 DAN SIMON P. AQUINODocument47 pagesScience11 - Q2 - Mod1 - GeneralBiology1 DAN SIMON P. AQUINOLyka Mae BenitoNo ratings yet
- Urea Plant Material Balance ACES Process PDFDocument7 pagesUrea Plant Material Balance ACES Process PDFArielSandovalNo ratings yet
- The Purple Sweet Potato Body Scrub Cream FormulatiDocument6 pagesThe Purple Sweet Potato Body Scrub Cream FormulatiniaNo ratings yet
- Jotun SteelmasterDocument13 pagesJotun SteelmasterKishan PradhanNo ratings yet
- Inorganica Chimica Acta: Research PaperDocument14 pagesInorganica Chimica Acta: Research PaperRuan ReisNo ratings yet
- History of LipstickDocument12 pagesHistory of Lipstickapi-368911467100% (1)
- Bioprinting and Its Applications in Tissue EngineeringDocument59 pagesBioprinting and Its Applications in Tissue EngineeringrezinhaakemiNo ratings yet
- Brochure Pt-Overview 2019 WebDocument13 pagesBrochure Pt-Overview 2019 WebPeter PandaNo ratings yet
- Case Study No.4 The Telltale Heart: Group 2 Nuñez, Refuerzo, Abalos, Almonte, AlmueteDocument11 pagesCase Study No.4 The Telltale Heart: Group 2 Nuñez, Refuerzo, Abalos, Almonte, AlmueteRejeanne MonroyNo ratings yet
- General Organic Chemistry - Iii: Section (A) : Solvents, Reagents and Leaving GroupsDocument20 pagesGeneral Organic Chemistry - Iii: Section (A) : Solvents, Reagents and Leaving GroupsGOURISH AGRAWALNo ratings yet
- Research PaperDocument6 pagesResearch PaperDhawalSoniNo ratings yet