You might also like
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (121)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (588)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (400)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5794)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1090)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2259)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (345)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (895)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (74)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- Parker HPD Product Bulletin (HY28-2673-01)Document162 pagesParker HPD Product Bulletin (HY28-2673-01)helden50229881No ratings yet
- Astm D 1196 PDFDocument3 pagesAstm D 1196 PDFSetyawan Chill Gates0% (1)
- Chapter 24 - The Solar SystemDocument36 pagesChapter 24 - The Solar SystemHeather Blackwell100% (1)
- P66 M10 CAT B Forms and Docs 04 10Document68 pagesP66 M10 CAT B Forms and Docs 04 10VinayNo ratings yet
- ProbDocument10 pagesProbKashif JawaidNo ratings yet
- Beautiful SpotsDocument2 pagesBeautiful SpotsLouise Yongco100% (1)
- Medabots-Rokusho Version (European) - Medal Codes (Part 1) (GBA Cheats) - CodeTwink ForumsDocument5 pagesMedabots-Rokusho Version (European) - Medal Codes (Part 1) (GBA Cheats) - CodeTwink Forumsdegraded 4resterNo ratings yet
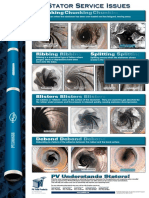
- Chunking Chunking Chunking: Stator Service IssuesDocument1 pageChunking Chunking Chunking: Stator Service IssuesGina Vanessa Quintero CruzNo ratings yet
- Nyamango Site Meeting 9 ReportDocument18 pagesNyamango Site Meeting 9 ReportMbayo David GodfreyNo ratings yet
- Trading Journal TDA Branded.v3.5 - W - Total - Transaction - Cost - BlankDocument49 pagesTrading Journal TDA Branded.v3.5 - W - Total - Transaction - Cost - BlankChristyann LojaNo ratings yet
- What Are The Advantages and Disadvantages of UsingDocument4 pagesWhat Are The Advantages and Disadvantages of UsingJofet Mendiola88% (8)
- Friday 25 Mar 12:15 AM Friday 25 Mar 5:30 AM: Emirates CGK DXBDocument3 pagesFriday 25 Mar 12:15 AM Friday 25 Mar 5:30 AM: Emirates CGK DXBDONI ARTANo ratings yet
- 3g Node B On Ip MediaDocument79 pages3g Node B On Ip MediaBsskkd KkdNo ratings yet
- Comparitive Study of Fifty Cases of Open Pyelolithotomy and Ureterolithotomy With or Without Double J Stent InsertionDocument4 pagesComparitive Study of Fifty Cases of Open Pyelolithotomy and Ureterolithotomy With or Without Double J Stent InsertionSuril VithalaniNo ratings yet
- Channel & Lomolino 2000 Ranges and ExtinctionDocument3 pagesChannel & Lomolino 2000 Ranges and ExtinctionKellyta RodriguezNo ratings yet
- Thermodynamic c106Document120 pagesThermodynamic c106Драгослав БјелицаNo ratings yet
- Abbas Ali Mandviwala 200640147: Ba1530: Information Systems and Organization StudiesDocument11 pagesAbbas Ali Mandviwala 200640147: Ba1530: Information Systems and Organization Studiesshayan sohailNo ratings yet
- A Brief Tutorial On Interval Type-2 Fuzzy Sets and SystemsDocument10 pagesA Brief Tutorial On Interval Type-2 Fuzzy Sets and SystemstarekeeeNo ratings yet
- ASHRAE Elearning Course List - Order FormDocument4 pagesASHRAE Elearning Course List - Order Formsaquib715No ratings yet
- Nse 2Document5 pagesNse 2dhaval gohelNo ratings yet
- Ethiopian Airlines-ResultsDocument1 pageEthiopian Airlines-Resultsabdirahmanguray46No ratings yet
- List of HTML TagsDocument5 pagesList of HTML TagsdorinadidNo ratings yet
- Rs2-Seamanship (Inc Anchoring, Mooring, Berthing, Pilot Ladder)Document19 pagesRs2-Seamanship (Inc Anchoring, Mooring, Berthing, Pilot Ladder)Mdpn. Salvador67% (3)
- Aspek Perpajakan Dalam Transfer Pricing: Related PapersDocument15 pagesAspek Perpajakan Dalam Transfer Pricing: Related PapersHasrawati AzisNo ratings yet
- Smart Plug Installation GuideDocument9 pagesSmart Plug Installation GuideFrancisco GuerreroNo ratings yet
- Garments Costing Sheet of LADIES Skinny DenimsDocument1 pageGarments Costing Sheet of LADIES Skinny DenimsDebopriya SahaNo ratings yet
- Module 2 TechnologyDocument20 pagesModule 2 Technologybenitez1No ratings yet
- Strategic Capital Management: Group - 4 Jahnvi Jethanandini Shreyasi Halder Siddhartha Bayye Sweta SarojDocument5 pagesStrategic Capital Management: Group - 4 Jahnvi Jethanandini Shreyasi Halder Siddhartha Bayye Sweta SarojSwetaSarojNo ratings yet
- Zigbee Technology:19-3-2010: Seminor Title DateDocument21 pagesZigbee Technology:19-3-2010: Seminor Title Dateitdep_gpcet7225No ratings yet
- SAP HR - Legacy System Migration Workbench (LSMW)Document5 pagesSAP HR - Legacy System Migration Workbench (LSMW)Bharathk KldNo ratings yet