You might also like
- Chronic Low Back Pain: Exploring Trends and Potential PredictorsDocument95 pagesChronic Low Back Pain: Exploring Trends and Potential Predictorssarranico4No ratings yet
- EquanimityDocument1 pageEquanimitysarranico4No ratings yet
- SC Germer ChapterDocument27 pagesSC Germer Chaptersarranico4No ratings yet
- Darwin and CompassionDocument2 pagesDarwin and Compassionsarranico4No ratings yet
- Chemical PhysicsDocument2 pagesChemical Physicssarranico4No ratings yet
- Mindfulness-Based Interventions in Schools-A Systematic Review and Meta-AnalysisDocument20 pagesMindfulness-Based Interventions in Schools-A Systematic Review and Meta-Analysissarranico4No ratings yet
- Mindfulness and Well-BeingDocument17 pagesMindfulness and Well-Beingsarranico4No ratings yet
- EquanimityDocument53 pagesEquanimitysarranico4No ratings yet
- How Does Mindfulness Meditation WorkDocument24 pagesHow Does Mindfulness Meditation Worksarranico4No ratings yet
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5794)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (890)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (399)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (344)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (587)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (73)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (265)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1090)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2219)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (119)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- Lear Jonathan Aristotelian InfinityDocument25 pagesLear Jonathan Aristotelian Infinityαλεξάνδρα σούσουNo ratings yet
- Lesson Plan Evaluation Five Macbeth Actvi December 5th 2Document2 pagesLesson Plan Evaluation Five Macbeth Actvi December 5th 2api-242677902No ratings yet
- Week 29 Year 1Document3 pagesWeek 29 Year 1Cikgu SyabilNo ratings yet
- Contoh Extended AbstractDocument8 pagesContoh Extended Abstractbintangabadi_techNo ratings yet
- Qualitative.31400 68960 1 SMDocument11 pagesQualitative.31400 68960 1 SMSyrel jane GarayNo ratings yet
- Lab Report RubricDocument2 pagesLab Report RubricjacnauNo ratings yet
- REHABILITATION OF MENTALLY ILL (Final)Document47 pagesREHABILITATION OF MENTALLY ILL (Final)gopikaNo ratings yet
- NT TeamworkDocument18 pagesNT TeamworkTaim SalahNo ratings yet
- TAG Common Grammar Mistakes With ExplanationsDocument43 pagesTAG Common Grammar Mistakes With ExplanationsMhel Ryan FloresNo ratings yet
- Sternberg - A Balance Theory of WisdomDocument19 pagesSternberg - A Balance Theory of WisdomSarah SiddiqiNo ratings yet
- A.2.2. Assessment CriteriaDocument2 pagesA.2.2. Assessment CriteriaAJ Masaga NamNo ratings yet
- Implicature in Panji Koming ComicDocument19 pagesImplicature in Panji Koming ComicMayla FPNo ratings yet
- Take a Position: Crafting an Argumentative Essay IntroductionDocument2 pagesTake a Position: Crafting an Argumentative Essay IntroductionhomamunfatNo ratings yet
- Trauma On Brain DevelopmentDocument29 pagesTrauma On Brain DevelopmentChirra Williams100% (1)
- Conditional PropositionsDocument8 pagesConditional Propositionsedniel maratasNo ratings yet
- Introduction and Review: EspañolDocument14 pagesIntroduction and Review: EspañolAlmyr Jay JimenezNo ratings yet
- Artigo Nervo Vago e Interocepção 22-11Document17 pagesArtigo Nervo Vago e Interocepção 22-11ThaisMaria DiasNo ratings yet
- Resolve Conflicts Using Negotiation & MediationDocument16 pagesResolve Conflicts Using Negotiation & MediationKrishnareddy KurnoolNo ratings yet
- A Critical History of Western PhilosophyDocument7 pagesA Critical History of Western PhilosophymahmudNo ratings yet
- Dll-Oral Communication Q1-W2Document5 pagesDll-Oral Communication Q1-W2Marie JavierNo ratings yet
- Word OrderDocument15 pagesWord OrderIndah EftanastariniNo ratings yet
- Needs Analysis in A CLIL Context: A Transfer From ESP: Miguel F. Ruiz-Garrido and Inmaculada Fortanet-GómezDocument10 pagesNeeds Analysis in A CLIL Context: A Transfer From ESP: Miguel F. Ruiz-Garrido and Inmaculada Fortanet-GómezKalvin KasongoNo ratings yet
- Varieties of PositioningDocument15 pagesVarieties of PositioningbendevaneNo ratings yet
- Training Needs AnalysisDocument9 pagesTraining Needs Analysiskavitha155No ratings yet
- Dependency in Linguistic Description, de Igor Mel'CukDocument114 pagesDependency in Linguistic Description, de Igor Mel'CukMarcelo Silveira100% (1)
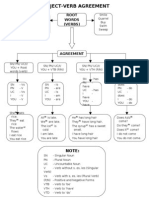
- Subject-Verb Agreement RulesDocument1 pageSubject-Verb Agreement RulesMohd AsyrafNo ratings yet
- Lip Reading Using CNN and LTSMDocument9 pagesLip Reading Using CNN and LTSMAlexandraZapucNo ratings yet
- Digital Unit Plan Template UpdatedDocument3 pagesDigital Unit Plan Template Updatedapi-398431038No ratings yet
- Italian 101Document14 pagesItalian 101Jamila Colleen M. BrionesNo ratings yet
- Year 4 Daily Lesson Plans: Skills Pedagogy (Strategy/Activity)Document3 pagesYear 4 Daily Lesson Plans: Skills Pedagogy (Strategy/Activity)Fatin Nur HafizahNo ratings yet