You might also like
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- ASTM Methode D 1066 Vapeur EchantillonnageDocument7 pagesASTM Methode D 1066 Vapeur Echantillonnageaaro_oraalNo ratings yet
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (587)
- D 3373 - 93 r98 - RdmznzmtukveDocument6 pagesD 3373 - 93 r98 - Rdmznzmtukvejamaljamal20No ratings yet
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (894)
- The Sum of The First N Natural Numbers - 9mathDocument3 pagesThe Sum of The First N Natural Numbers - 9mathjamaljamal20No ratings yet
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- D 1688 - 02 - Rde2odgDocument9 pagesD 1688 - 02 - Rde2odgjamaljamal20No ratings yet
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (399)
- Reporting Results of Examination and Analysis of Water-Formed DepositsDocument3 pagesReporting Results of Examination and Analysis of Water-Formed DepositsphaindikaNo ratings yet
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (73)
- D 596 - 01 - Rdu5ngDocument6 pagesD 596 - 01 - Rdu5ngjamaljamal20No ratings yet
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5794)
- D 932 - 85 R02 - RDKZMGDocument8 pagesD 932 - 85 R02 - RDKZMGjamaljamal20No ratings yet
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- ASTM Methode D 1066 Vapeur EchantillonnageDocument7 pagesASTM Methode D 1066 Vapeur Echantillonnageaaro_oraalNo ratings yet
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- D 934 - 80 R03 - RdkznaDocument9 pagesD 934 - 80 R03 - Rdkznajamaljamal20No ratings yet
- Astm D 513 - 02Document8 pagesAstm D 513 - 02phaindikaNo ratings yet
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- D 858 - 02 - Rdg1oaDocument8 pagesD 858 - 02 - Rdg1oajamaljamal20No ratings yet
- D 3557 - 95 - Rdm1ntctukveDocument18 pagesD 3557 - 95 - Rdm1ntctukvejamaljamal20No ratings yet
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- D 888 - 03 - Rdg4oaDocument9 pagesD 888 - 03 - Rdg4oajamaljamal20No ratings yet
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1090)
- D 807 - 00 - RDGWNWDocument6 pagesD 807 - 00 - RDGWNWjamaljamal20No ratings yet
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2219)
- D 857 - 95 - Rdg1ny05nqDocument4 pagesD 857 - 95 - Rdg1ny05nqjamaljamal20No ratings yet
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- Elements in Water by Direct-Current Plasma Atomic Emission SpectrosDocument7 pagesElements in Water by Direct-Current Plasma Atomic Emission Spectrosjamaljamal20No ratings yet
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (344)
- D 2077 - 92 R98 - RdiwnzcDocument1 pageD 2077 - 92 R98 - Rdiwnzcjamaljamal20No ratings yet
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (265)
- D 3263 - 82 r99 - RdmynjmtodjsotlfmqDocument8 pagesD 3263 - 82 r99 - Rdmynjmtodjsotlfmqjamaljamal20No ratings yet
- D 2527 - 83 R01 - Rdi1mjcDocument2 pagesD 2527 - 83 R01 - Rdi1mjcjamaljamal20No ratings yet
- D 2433 - 93 r98 - Rdi0mzmtotnsotgDocument6 pagesD 2433 - 93 r98 - Rdi0mzmtotnsotgjamaljamal20No ratings yet
- Aluminum in Water D 857 - 02 RDG1NW - PDFDocument5 pagesAluminum in Water D 857 - 02 RDG1NW - PDFstevgonNo ratings yet
- D 888 - 03 - Rdg4oaDocument9 pagesD 888 - 03 - Rdg4oajamaljamal20No ratings yet
- D 2777 - 98 - Rdi3nzctotgDocument15 pagesD 2777 - 98 - Rdi3nzctotgjamaljamal20No ratings yet
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- F 148 - 95 - Rje0oc05nqDocument2 pagesF 148 - 95 - Rje0oc05nqjamaljamal20No ratings yet
- D 928 - 03 - Rdkyoc1sruqDocument3 pagesD 928 - 03 - Rdkyoc1sruqjamaljamal20No ratings yet
- D 1331 - 89 R01 - RdezmzeDocument3 pagesD 1331 - 89 R01 - Rdezmzejamaljamal20No ratings yet
- D 726 - 94 R99 - Rdcyni05nfi5oqDocument3 pagesD 726 - 94 R99 - Rdcyni05nfi5oqjamaljamal20No ratings yet
- Surface Wettability of Paper (Angle-of-Contact Method) : Standard Test Method ForDocument4 pagesSurface Wettability of Paper (Angle-of-Contact Method) : Standard Test Method Forjamaljamal20No ratings yet
- D 5077 - 90 r97 - RduwnzctotbsotcDocument2 pagesD 5077 - 90 r97 - Rduwnzctotbsotcjamaljamal20No ratings yet
- Concentrated Impacts To Transport Packages: Standard Test Method ForDocument4 pagesConcentrated Impacts To Transport Packages: Standard Test Method Forjamaljamal20100% (1)
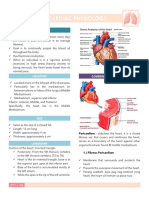
- The Heart: Structure and FunctionDocument2 pagesThe Heart: Structure and FunctionHolly May MontejoNo ratings yet
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (119)
- Establishment of Cacao Nursery and ManagementDocument121 pagesEstablishment of Cacao Nursery and ManagementPlantacion de Sikwate100% (29)
- Pro67-A-13 Prep of McFarland StdsDocument6 pagesPro67-A-13 Prep of McFarland StdsWisnu Ary NugrohoNo ratings yet
- Non-Critically Ill Hyperglycemia ManagementDocument39 pagesNon-Critically Ill Hyperglycemia ManagementHerry KongkoNo ratings yet
- Heart beats 100,000 times dailyDocument7 pagesHeart beats 100,000 times dailyKyle MañiboNo ratings yet
- Molecular Drug Targets-1Document75 pagesMolecular Drug Targets-1Phoebe Llamelo100% (1)
- LAS 1 Cell TheoryDocument12 pagesLAS 1 Cell TheoryJeremie CataggatanNo ratings yet
- Organizational Behavior Field Map and Perception FactorsDocument32 pagesOrganizational Behavior Field Map and Perception FactorsPratik TagwaleNo ratings yet
- Cucumber Seed Germination: Effect and After-Effect of Temperature TreatmentsDocument7 pagesCucumber Seed Germination: Effect and After-Effect of Temperature Treatmentsprotap uddin PolenNo ratings yet
- Composite Materials For Biomedical Applications: A ReviewDocument17 pagesComposite Materials For Biomedical Applications: A ReviewDuc Anh NguyenNo ratings yet
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- An Efficient Micropropagation Protocol For Eclipta AlbaDocument6 pagesAn Efficient Micropropagation Protocol For Eclipta AlbaSuprabuddha KunduNo ratings yet
- Psychiatric InterviewDocument54 pagesPsychiatric InterviewzarrarNo ratings yet
- Atheena Milagi Pandian. SDocument3 pagesAtheena Milagi Pandian. SAtheena PandianNo ratings yet
- Wbtet 2012Document7 pagesWbtet 2012Partha BatabyalNo ratings yet
- Coccon Jacket Pattern A4 PDFDocument16 pagesCoccon Jacket Pattern A4 PDFdaisy22No ratings yet
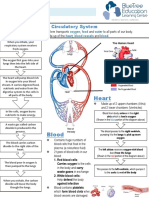
- Circulatory System Notes SLDocument1 pageCirculatory System Notes SLRathna PalaniappanNo ratings yet
- Quiz - Mosndarja e Kromozomeve Gjate MejozwsDocument5 pagesQuiz - Mosndarja e Kromozomeve Gjate MejozwsleartaNo ratings yet
- Mapping Aqsha 1: Heart Failure, Esophageal Cancer, and MoreDocument7 pagesMapping Aqsha 1: Heart Failure, Esophageal Cancer, and MorePutri Rahmadhani Ngakpaniklage AsdsNo ratings yet
- Thymoma: Rawa MuhsinDocument53 pagesThymoma: Rawa MuhsinRawa MuhsinNo ratings yet
- Material Properties of Poly (Propylene Carbonates)Document21 pagesMaterial Properties of Poly (Propylene Carbonates)Adrian Fernandez BelloNo ratings yet
- Lca Mangatarem1 PDFDocument58 pagesLca Mangatarem1 PDFBea 'Jane0% (1)
- PhysioEx Exercise 1 Activity 2Document3 pagesPhysioEx Exercise 1 Activity 2CLAUDIA ELISABET BECERRA GONZALESNo ratings yet
- Chapter 1 of Blue MindDocument23 pagesChapter 1 of Blue MindSouthern California Public Radio100% (1)
- EcotourismDocument9 pagesEcotourismyoshoworNo ratings yet
- Udc Vol 1Document921 pagesUdc Vol 1Dani TrashNo ratings yet
- BS 08545-2013Document97 pagesBS 08545-2013Stanislav PerevezentsevNo ratings yet
- SWP 331 Final Exam NotesDocument27 pagesSWP 331 Final Exam Notesdcadenas1No ratings yet
- Dr. Morse's Herbal Formulation ListDocument10 pagesDr. Morse's Herbal Formulation ListTom50% (2)
- Seroepidemiology of Clamaydia TDocument8 pagesSeroepidemiology of Clamaydia TMulatuNo ratings yet
- To Study The Quantity Case in Present in Different Sample of MilkDocument7 pagesTo Study The Quantity Case in Present in Different Sample of Milkbelly4u100% (2)
- Trevor Kletz Compendium: His Process Safety Wisdom Updated for a New GenerationFrom EverandTrevor Kletz Compendium: His Process Safety Wisdom Updated for a New GenerationNo ratings yet
- Guidelines for Vapor Cloud Explosion, Pressure Vessel Burst, BLEVE, and Flash Fire HazardsFrom EverandGuidelines for Vapor Cloud Explosion, Pressure Vessel Burst, BLEVE, and Flash Fire HazardsNo ratings yet
- Functional Safety from Scratch: A Practical Guide to Process Industry ApplicationsFrom EverandFunctional Safety from Scratch: A Practical Guide to Process Industry ApplicationsNo ratings yet
- Perfume Engineering: Design, Performance and ClassificationFrom EverandPerfume Engineering: Design, Performance and ClassificationRating: 4 out of 5 stars4/5 (5)
- Piping and Pipeline Calculations Manual: Construction, Design Fabrication and ExaminationFrom EverandPiping and Pipeline Calculations Manual: Construction, Design Fabrication and ExaminationRating: 4 out of 5 stars4/5 (18)
- Nuclear Energy in the 21st Century: World Nuclear University PressFrom EverandNuclear Energy in the 21st Century: World Nuclear University PressRating: 4.5 out of 5 stars4.5/5 (3)