You might also like
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5794)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1090)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- Essential Calculus Skills Practice Workbook With Full SolutionsDocument528 pagesEssential Calculus Skills Practice Workbook With Full SolutionsGerardo Navarro Sánchez94% (65)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (895)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (588)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (344)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (120)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (399)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2259)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- Joby Aviation - Analyst Day PresentationDocument100 pagesJoby Aviation - Analyst Day PresentationIan TanNo ratings yet
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (73)
- Book Index The Art of Heavy TransportDocument6 pagesBook Index The Art of Heavy TransportHermon Pakpahan50% (2)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- RD Sharma Class8 SolutionsDocument2 pagesRD Sharma Class8 Solutionsncertsoluitons100% (2)
- Chapter 1 TransportDocument1 pageChapter 1 TransportFatin MinhatNo ratings yet
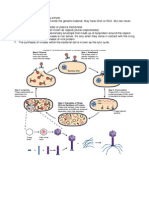
- Biodiversity VirusesDocument4 pagesBiodiversity VirusesFatin MinhatNo ratings yet
- PhotosynthesisDocument1 pagePhotosynthesisFatin MinhatNo ratings yet
- The Human Vertebral ColumnDocument1 pageThe Human Vertebral ColumnFatin MinhatNo ratings yet
- 5Document1 page5Fatin MinhatNo ratings yet
- Microorganisms SC f5Document3 pagesMicroorganisms SC f5Fatin MinhatNo ratings yet
- Species AbundancesDocument3 pagesSpecies AbundancesFatin MinhatNo ratings yet
- Template Business CardDocument1 pageTemplate Business CardFatin MinhatNo ratings yet
- 2Document1 page2Fatin MinhatNo ratings yet
- Herbal Does Not Mean Innocuous: Ten Cases of Severe Hepatotoxicity Associated With Dietary Supplements From HerbalifeÒ ProductsqDocument6 pagesHerbal Does Not Mean Innocuous: Ten Cases of Severe Hepatotoxicity Associated With Dietary Supplements From HerbalifeÒ ProductsqMohammed AyubNo ratings yet
- The South Luconia ShoalsDocument1 pageThe South Luconia ShoalsFatin MinhatNo ratings yet
- January 2012: Lam Wah Ee Along & Abah (Appmnt)Document12 pagesJanuary 2012: Lam Wah Ee Along & Abah (Appmnt)Fatin MinhatNo ratings yet
- FitnessDocument1 pageFitnessFatin MinhatNo ratings yet
- Book 123Document2 pagesBook 123Fatin MinhatNo ratings yet
- Answers q1Document9 pagesAnswers q1Kavitha RajendranNo ratings yet
- Ayat Dan DoaDocument2 pagesAyat Dan DoaFatin MinhatNo ratings yet
- Q2Document60 pagesQ2Fatin MinhatNo ratings yet
- Test 8 D - Unit 2Document3 pagesTest 8 D - Unit 2IONELA MIHAELA POPANo ratings yet
- Tabla9 1Document1 pageTabla9 1everquinNo ratings yet
- Designing and Building A Computer TableDocument9 pagesDesigning and Building A Computer Tablemaster_codersNo ratings yet
- Critical Analysis of W.H Auden Poems The PDFDocument8 pagesCritical Analysis of W.H Auden Poems The PDFMöĤämmĔd äĹ-ŚäÁdï100% (1)
- Maintenance Páginas-509-580Document72 pagesMaintenance Páginas-509-580Alexandra Gabriela Pacheco PrietoNo ratings yet
- Water Filling MachineDocument15 pagesWater Filling Machinepallab D RozarioNo ratings yet
- Metaphysics of LucretiusDocument6 pagesMetaphysics of LucretiusChristopher BennettNo ratings yet
- Esterification Oil of WintergreenDocument8 pagesEsterification Oil of WintergreenMaria MahusayNo ratings yet
- 3397 - Ciat LDC 300VDocument71 pages3397 - Ciat LDC 300VPeradNo ratings yet
- Đề 17Document11 pagesĐề 17Nguyen CuongNo ratings yet
- Regression Analysis Random Motors ProjectDocument22 pagesRegression Analysis Random Motors ProjectPrateek AgrawalNo ratings yet
- DIVAR IP All-In-One 7000 3U Datasheet 51 en 66297110155Document5 pagesDIVAR IP All-In-One 7000 3U Datasheet 51 en 66297110155Javier RochaNo ratings yet
- Patel 2013Document116 pagesPatel 2013hnphuocNo ratings yet
- Chapter 7: Protein Function Part I: Myoglobin and HemoglobinDocument27 pagesChapter 7: Protein Function Part I: Myoglobin and HemoglobineliNo ratings yet
- Ap, Lrrsisal of Roentgenograph, Ic: I SsayDocument30 pagesAp, Lrrsisal of Roentgenograph, Ic: I SsayMindaugasStacevičiusNo ratings yet
- Climbing FormworkDocument4 pagesClimbing FormworkAshwin B S RaoNo ratings yet
- Recruitment and Selection in Canada 7Th by Catano Wiesner Full ChapterDocument22 pagesRecruitment and Selection in Canada 7Th by Catano Wiesner Full Chaptermary.jauregui841100% (51)
- Armadio Presentation-2019Document45 pagesArmadio Presentation-2019Subhash Singh TomarNo ratings yet
- CCNA Training New CCNA - RSTPDocument7 pagesCCNA Training New CCNA - RSTPokotete evidenceNo ratings yet
- Minimalist KWL Graphic OrganizerDocument2 pagesMinimalist KWL Graphic OrganizerIrish Nicole AlanoNo ratings yet
- Reynold A. Nicholson - The Mystics of IslamDocument65 pagesReynold A. Nicholson - The Mystics of IslamLuminon SamanNo ratings yet
- Notes Transfer of Thermal EnergyDocument12 pagesNotes Transfer of Thermal Energymahrosh mamoon100% (2)
- GBJ0232 - en GLX 3101 T2Document43 pagesGBJ0232 - en GLX 3101 T2mnbvqwert100% (2)
- Islam and Civilization (Analysis Study On The History of Civilization in Islam) Muhammad Hifdil IslamDocument18 pagesIslam and Civilization (Analysis Study On The History of Civilization in Islam) Muhammad Hifdil IslamLoveth KonniaNo ratings yet
- Welcome To Our 2Nd Topic: History of VolleyballDocument6 pagesWelcome To Our 2Nd Topic: History of VolleyballDharyn KhaiNo ratings yet
- CulvertsDocument18 pagesCulvertsAmmar A. Ali100% (1)