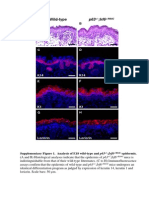
You might also like
- Babylonian Orion CetusDocument4 pagesBabylonian Orion CetusmardukNo ratings yet
- Olmec ArtDocument246 pagesOlmec Artmarduk100% (7)
- Handbook of the Aging BrainFrom EverandHandbook of the Aging BrainEugenia WangNo ratings yet
- Mutation and Polymorphism Analysis of The Human Homogentisate 1,2-Dioxygenase Gene in Alkaptonuria PatientsDocument9 pagesMutation and Polymorphism Analysis of The Human Homogentisate 1,2-Dioxygenase Gene in Alkaptonuria PatientscristinaNo ratings yet
- Brief Report: Correction of A Genetic Disease in Mouse Via Use of CRISPR-Cas9Document4 pagesBrief Report: Correction of A Genetic Disease in Mouse Via Use of CRISPR-Cas9Aida KNo ratings yet
- JHG 201178Document3 pagesJHG 201178DAVID TERRONESNo ratings yet
- SCARDUSconf.ffffinalDocument6 pagesSCARDUSconf.ffffinalbucajselma20No ratings yet
- pnas.202001270Document10 pagespnas.202001270diego.battiatoNo ratings yet
- 10 1016@j Gene 2019 04 080Document8 pages10 1016@j Gene 2019 04 080Gowtham ShanNo ratings yet
- Genetic Polymorphisms Adjacent To The CAG Repeat Influence Clinical Features at Onset in Huntington's DiseaseDocument5 pagesGenetic Polymorphisms Adjacent To The CAG Repeat Influence Clinical Features at Onset in Huntington's DiseaseShavana RajkumarNo ratings yet
- jcinvest00095-0208Document7 pagesjcinvest00095-0208bilo47762No ratings yet
- Acfrogc0kyj2l3 A23iobc O3pbzxu4d4z2 Bov Kj7jsvkubumlfopnv3ljry6fpadaicfkrigsyg3hhae3ssru3qgalzr Enevkfui208ghy23p0kmzsgejh Fleoq2gik8xm4bzstlxdt8t02Document9 pagesAcfrogc0kyj2l3 A23iobc O3pbzxu4d4z2 Bov Kj7jsvkubumlfopnv3ljry6fpadaicfkrigsyg3hhae3ssru3qgalzr Enevkfui208ghy23p0kmzsgejh Fleoq2gik8xm4bzstlxdt8t02Marco TúlioNo ratings yet
- Hutchinson-Gilford Progeria Syndrome: Mini ReviewDocument7 pagesHutchinson-Gilford Progeria Syndrome: Mini ReviewBrenda Argomedo CamposNo ratings yet
- Janetopoulos Science2001Document5 pagesJanetopoulos Science2001Psych PsychhNo ratings yet
- Tian 2012Document16 pagesTian 2012Karina Caudillo GamezNo ratings yet
- MCLR Studies in Dogs With Melanistic Mask or Brindle PatternsDocument5 pagesMCLR Studies in Dogs With Melanistic Mask or Brindle PatternsSmith SanNo ratings yet
- CAG Repeat Instability at SCA2 Locus - Anchoring CAA Interruptions and Linked Single Nucleotide PolymorphismsDocument10 pagesCAG Repeat Instability at SCA2 Locus - Anchoring CAA Interruptions and Linked Single Nucleotide PolymorphismsSarahNo ratings yet
- Association of Neuregulin 1 With Schizophrenia in SECOND COHORT From The Scottish Population. 2003.Document5 pagesAssociation of Neuregulin 1 With Schizophrenia in SECOND COHORT From The Scottish Population. 2003.Fahrunnisa NurdinNo ratings yet
- Bahanrefohmori 2001Document4 pagesBahanrefohmori 2001Fitri anaNo ratings yet
- Vertical Starch-Gel IN Some Rheumatic Diseases : ElectrophoresisDocument4 pagesVertical Starch-Gel IN Some Rheumatic Diseases : ElectrophoresisWiwik yuliatiNo ratings yet
- Morphological and Physiological Interactions of NG2-glia With Astrocytes and NeuronsDocument10 pagesMorphological and Physiological Interactions of NG2-glia With Astrocytes and NeuronsRaffaello MobiliaNo ratings yet
- GBAmutations in Gaucher Type I Venezuelan Patients - Ethnic Originsand FrequenciesDocument7 pagesGBAmutations in Gaucher Type I Venezuelan Patients - Ethnic Originsand FrequenciesGilberto GomezNo ratings yet
- PubmedDocument7 pagesPubmedcristinaNo ratings yet
- Takeuchi 2000Document4 pagesTakeuchi 2000dad dzd adaNo ratings yet
- Comparative pharmacology of adrenergic α2C receptors coupled to Ca2+ signaling through different Gα proteinsDocument9 pagesComparative pharmacology of adrenergic α2C receptors coupled to Ca2+ signaling through different Gα proteinsChasconaNo ratings yet
- The Genetic Aspect and Morphological Appearance of AchondrogenesisDocument10 pagesThe Genetic Aspect and Morphological Appearance of AchondrogenesisRizka RamadaniNo ratings yet
- Unilateral Dried Out Eyesight SyndromeisjmvDocument1 pageUnilateral Dried Out Eyesight Syndromeisjmvclassnephew5No ratings yet
- 15-Deoxy - Prostaglandin J Induces IL-8 and GM-CSF in A Human Airway Epithelial Cell Line (NCI-H)Document7 pages15-Deoxy - Prostaglandin J Induces IL-8 and GM-CSF in A Human Airway Epithelial Cell Line (NCI-H)Roy GoldenNo ratings yet
- s41598 022 25914 8Document13 pagess41598 022 25914 8AllyNo ratings yet
- Recurrent de Novo Point Mutations in Lamin A Cause Hutchinson-Gilford Progeria SyndromeDocument6 pagesRecurrent de Novo Point Mutations in Lamin A Cause Hutchinson-Gilford Progeria SyndromeGho VinsenNo ratings yet
- Mutations in VKORC1 Cause Warfarin Resistance and Multiple Coagulation Factor Deficiency Type 2Document5 pagesMutations in VKORC1 Cause Warfarin Resistance and Multiple Coagulation Factor Deficiency Type 2Eva SefianaNo ratings yet
- Wild-Type: p63 Irf6Document5 pagesWild-Type: p63 Irf6Nadya PurwantyNo ratings yet
- Gly Paper 1Document15 pagesGly Paper 1drirfankhan69No ratings yet
- Antibody GAPDHDocument3 pagesAntibody GAPDHWendy Aide Castro MoraNo ratings yet
- Duplication Source Place Might Contribute To Hereditary Variation in Trypanosoma CruziwlcwzDocument1 pageDuplication Source Place Might Contribute To Hereditary Variation in Trypanosoma Cruziwlcwzgermanmallet1No ratings yet
- Detection of Single Nucleotide Polymorphism of A1/A2 Variants of Beta Caseain Gene in Umbalachery Cattle by Tetra Arms PCRDocument7 pagesDetection of Single Nucleotide Polymorphism of A1/A2 Variants of Beta Caseain Gene in Umbalachery Cattle by Tetra Arms PCRIndian Journal of Veterinary and Animal Sciences RNo ratings yet
- Notes Identification of Bovine and Porcine Rotavirus G Types by PCRDocument3 pagesNotes Identification of Bovine and Porcine Rotavirus G Types by PCRBhaskar KarumuriNo ratings yet
- Brief Communications: Total Silencing by Intron-Spliced Hairpin RnasDocument2 pagesBrief Communications: Total Silencing by Intron-Spliced Hairpin RnasDiego Tegomas DomínguezNo ratings yet
- FISH-MLPA-Array Based Detection For 18q DeletionDocument8 pagesFISH-MLPA-Array Based Detection For 18q DeletionladybieibiNo ratings yet
- Oral Ambroxol Increases Brain Glucocerebrosidase Activity in A Nonhuman PrimateDocument3 pagesOral Ambroxol Increases Brain Glucocerebrosidase Activity in A Nonhuman PrimateTia MutiaraNo ratings yet
- Prasher - GFP StructureDocument5 pagesPrasher - GFP StructureArjun AggarwalNo ratings yet
- SUR - Frei Et Al.Document6 pagesSUR - Frei Et Al.Marcelo EndresNo ratings yet
- PpokDocument7 pagesPpokSkoy KuyNo ratings yet
- Articulo Polineuropatía 2Document1 pageArticulo Polineuropatía 2Paula JoyaNo ratings yet
- Expression of Neugc On Pig Corneas and Its Potential Significance in Pig Corneal XenotransplantationDocument9 pagesExpression of Neugc On Pig Corneas and Its Potential Significance in Pig Corneal XenotransplantationJoseph Chipana GutierrezNo ratings yet
- Animal Genetics 2022Document4 pagesAnimal Genetics 2022Kö TänyinNo ratings yet
- Suran To 1998Document6 pagesSuran To 1998an saidNo ratings yet
- 1 s2.0 S0306452216300665 MainDocument15 pages1 s2.0 S0306452216300665 MainPeter TuNo ratings yet
- 18 PDFDocument3 pages18 PDFjohn4jNo ratings yet
- tmp5C3D TMPDocument10 pagestmp5C3D TMPFrontiersNo ratings yet
- Shauni PosterDocument1 pageShauni Posterapi-268469441No ratings yet
- GuillainBarrE EVR CENETECDocument3 pagesGuillainBarrE EVR CENETECDavid BVNo ratings yet
- 4-Madhu Ygga 2004Document8 pages4-Madhu Ygga 2004AsmaNo ratings yet
- Molecular Survey of Babesia Infection in Dogs in Okinawa, Japan 2004 Veterinary ParasitologyDocument6 pagesMolecular Survey of Babesia Infection in Dogs in Okinawa, Japan 2004 Veterinary ParasitologyGabriela Victoria MartinescuNo ratings yet
- Choy 1996Document6 pagesChoy 1996Araceli Enríquez OvandoNo ratings yet
- 7554 13381 1 PB PDFDocument10 pages7554 13381 1 PB PDFRamla YaniNo ratings yet
- INTRODUCTION 1.1: Structure of Hemoglobin (PDFDrive)Document102 pagesINTRODUCTION 1.1: Structure of Hemoglobin (PDFDrive)Tizaa NassaNo ratings yet
- Radioimmunoassays For Glutamic Acid Decarboxylase (GAD65) and GAD65 Autoantibodies Using 35S or 3H Recombinant Human LigandsDocument11 pagesRadioimmunoassays For Glutamic Acid Decarboxylase (GAD65) and GAD65 Autoantibodies Using 35S or 3H Recombinant Human LigandsAudhy RaflesNo ratings yet
- 2415 1.tifDocument8 pages2415 1.tifchuck212No ratings yet
- Short Report A Genetic Variation of Echinococcus Canadendis (G7) in MexicoDocument5 pagesShort Report A Genetic Variation of Echinococcus Canadendis (G7) in MexicoAronyMartinezNo ratings yet
- 1 s2.0 S0042682206003928 MainDocument5 pages1 s2.0 S0042682206003928 MainAlberto VilladiegoNo ratings yet
- Reidla 2003 Haplogroup XDocument13 pagesReidla 2003 Haplogroup XmardukNo ratings yet
- Letters To The EditorDocument7 pagesLetters To The EditormardukNo ratings yet
- Malhi 2004 NA Mtdna DiversityDocument22 pagesMalhi 2004 NA Mtdna DiversitymardukNo ratings yet
- Mtdna NW NA EshlemanDocument21 pagesMtdna NW NA EshlemanmardukNo ratings yet
- N Vs (1) S MesopotaDocument6 pagesN Vs (1) S Mesopotamarduk100% (1)
- PyramidsDocument15 pagesPyramidsmardukNo ratings yet
- UssherDocument21 pagesUsshermardukNo ratings yet
- MtDNA Haplogroup X An Ancient Link Between Europe Western Asia and North AmericaDocument10 pagesMtDNA Haplogroup X An Ancient Link Between Europe Western Asia and North AmericaspanishvcuNo ratings yet
- IGCP Paper - Archaeology and Natural History Observations Concerning The Caspian Sea Ver1Document27 pagesIGCP Paper - Archaeology and Natural History Observations Concerning The Caspian Sea Ver1marduk100% (5)
- SubRosa Issue1-SpreadDocument26 pagesSubRosa Issue1-Spreadmarduk100% (4)
- NazcaDocument2 pagesNazcamarduk100% (1)
- CosmologyDocument5 pagesCosmologymardukNo ratings yet
- BibleDocument22 pagesBiblemardukNo ratings yet
- BulmerDocument22 pagesBulmermardukNo ratings yet
- 2007, The Great Pyramid Debate, 29th ICMADocument60 pages2007, The Great Pyramid Debate, 29th ICMAmardukNo ratings yet
- Genomics Refutes An Exclusively African Origin of HumansDocument56 pagesGenomics Refutes An Exclusively African Origin of HumansmardukNo ratings yet
- 000 A - Rituals Asl anguage-INTERESTINGDocument23 pages000 A - Rituals Asl anguage-INTERESTINGmardukNo ratings yet